Морфология пауков
Размеры пауков колеблются в очень широких пределах - от 0.8 мм (некоторые Oonopidae) до 90 мм (Theraphosa leblondi Latr.). Крупные формы среди пауков, вообще говоря, довольно обычны, в особенности в тропиках, однако у громадного большинства европейских видов длина не превышает 1-1.5 см. Эволюция отряда пауков в целом сопровождается заметным уменьшением размеров, подобно тому как это имеет место в классе насекомых (Insectа). Примитивные формы (например, представители подотрядов Liphistiomorphae и Mygalomorphae) имеют очень значительные размеры (порядка нескольких сантиметров). В то же время филогенетически молодые и прогрессивные группы, объединяющие огромное число видов, характеризуются весьма скромными размерами. Таковы весьма обычные в нашей фауне представители сем. Мicryphantidae, которые в немецкой литературе получили название "пауков-карликов" ("Zwergspinnen").
Форма тела пауков также заметно варьирует. Головогрудь в этом отношении оказывается менее эластичной, чем брюшко. Появление на головогруди горба, выроста, выступа и т. д. может быть связано либо с исключительным развитием и морфо-функциональной модификацией ядовитых желез (Scytodes thoracica Latr., см. рис. 63), либо с адаптацией к спариванию (подсем. Walckenaerinae из Micryphantidae). В последнем случае этот признак обнаруживается только у еамцрв. Возвышения, возникающие в области глазного поля, направлены, по-видимому, на увеличение площади обзора неподвижных глаз. Однако появление у Walckenaera стебелька с глазами, сидящими на его вершине (рис. 845, 847), вряд ли может быть объяснено с этой точки зрения. Брюшко у большинства пауков округлое, иногда почти сферическое, или яйцевидное. Некоторые замечательные примеры вариаций формы брюшка у тропических пауков показаны на рис. 1-4. Появление на брюшке различных выростов, шипов, бугорков и бородавок связано, возможно, с миметическими явлениями.
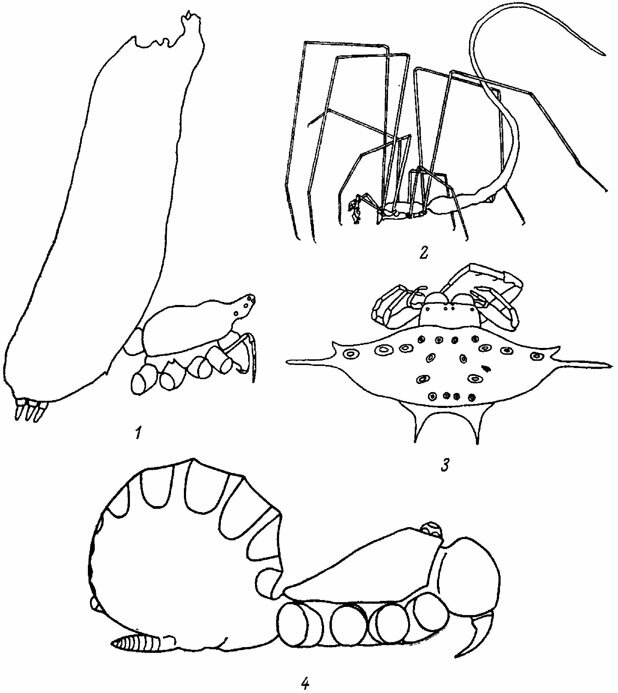
Рис. 1-4. Форма тела некоторых тропических пауков. По Милло. Рис. 1. Poltys columnaris Thor. (Araneidae). Рис. 2. Ariamnes helminthoides Sim. (Theridiidae), Рис. 3. Gasterocantha importuna Pick.-Cambr. (Araneidae). Рис. 4. Liphistius malayanus Abr. (Liphistiidae)
Довольно обычной модификацией формы тела у пауков, затрагивающей как головогрудь, так и брюшко, является продольное вытягивание тела, конвергентно развивающееся в самых различных семействах (Tetragnatha из Tetragnathidae, Tibellus из Thomisidae, Miagrammopes из Uloboridae, многие Рhо1cidae и др.). У ряда тропических видов тело в результате подобного вытягивания приобретает палочковидную или даже нитевидную форму (Ariamnes из Theridiidae, рис. 2). У видов, обитающих под корой или под камнями, наблюдается дорсо-вентральное сплющивание тела (Araneus umbraticus Сl. из Araneidae, Coriarachne и Oxyptila из Thomisidae). Ряд специфических изменений обычной формы тела связан с широко распространенным у пауков явлением имитации муравьев (рис. 5). В этом случае обычно наблюдается удлинение стебелька, а также появление перетяжек на головогруди и на брюшке (Peckham, 1889; Pocock, 1909; Dahl, 1913; Heikertinger, 1954; Wiehle, 1954). Известны также отдельные случаи подражания пауков жукам и осам-немкам (Dahl, 1903; Bristowe, 1939-1941).
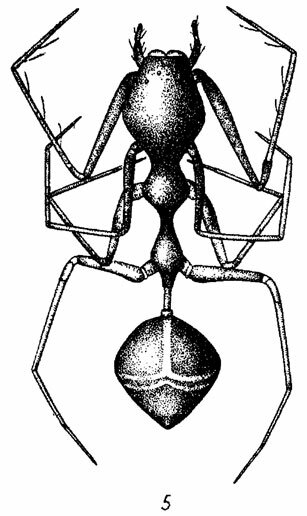
Рис. 5. Myrmecium gounellei Sim. (Clubionidae) - пример совершенной имитации муравья пауком. По Милло
Окраска пауков очень разнообразна. Хотя большинство видов окрашено в темные цвета - в черные, сероватые или коричневые, - некоторые тропические формы (Peucetia, Leucauge) имеют очень яркую окраску. Многие Salticidae и представители рода Micaria из Clubionidae имеют красивую металлически блестящую окраску, которая создается интерференцией света на чешуйках и чешуйчатых волосках. В зависимости от расположения чешуек и направления световых лучей на теле паука появляется зеленый, бронзовый, оранжевый, фиолетовый или красный отблеск. Остальные типы окраски пауков связаны с отложением пигментов в цитоплазме гиподермальных клеток, в гемолимфе, в кутикуле и в волосках (Millot, 1926).
Криптическая (покровительственная) окраска тела свойственна очень многим паукам и несомненно обеспечивает защиту этих животных от различных врагов (Dahl, 1905, 1913; Котт, 1950). Пауки, обитающие на коре, имеют бурую, коричневую или серую пятнистую окраску (Marpissa muscosa Cl., Philodromus fuscomarginatus de Geer). Живущие среди лишайников виды (например, Eustala anastera Walck. из Северной Америки) по своей окраске очень сильно напоминают лишайник, а виды, обитающие в траве (например, европейский Micrommata roseum Cl.), окрашены в зеленые цвета.
У встречающегося на цветках паука-крабохода (Misumena vatia Cl.) покровительственная окраска особенно эффективна, так как этот паук в зависимости от цвета окружающего субстрата может принимать белую, желтую, розовую или бледно-зеленую окраску (Packard, 1905; Kerville, 1907; Rabaud, 1918, 1919; Gabritschevsky, 1927). Некоторые крупные пауки имеют типичную предостерегающую окраску в виде сочетания черных и желтых полос (Argiope bruennichi Scop.).
В окраске разных полов иногда очень сильно проявляется половой диморфизм; самец паука обычно окрашен в более яркие и контрастные цвета. Такова, например, ярко-красная с черным расцветка брюшка у самцов Eresus niger Pet.
Головогрудь (cephalothorax) образуется в результате слияния акрона и всех сегментов просомы (сегмента хелицер, педипальп и 4 сегментов ходильных ног). У современных пауков следы сегментации головогруди полностью исчезли и имеющиеся на дорсальной стороне ее поперечные бороздки не отвечают границам сегментов.
Сверху головогрудь покрывается плотным хитиновым спинным щитом (саrарах), составленным из слившихся тергитов просомы (рис. 6). В задней части спинного щита обычно имеется небольшая медиальная ямка (fovea media), от которой отходят неглубокие радиальные бороздки. Первая пара радиальных бороздок отделяет переднюю часть спинного щита - голову от задней его части - груди. Голова, которая обычно слегка приподнята над грудью, несет глаза и хелицеры.
Глаза (ocelli), как и у других паукообразных, всегда простые. В типичном случае у пауков имеется 8 глаз. Эта цифра отвечает максимально возможному числу их и может уменьшаться до 6 (у Sicariidae, Dysderidae, Oonopidae, у некоторых Pholcidae), до 4 (у Tetrablemma) или даже до 2 (у Nops и Malta). Наконец, у типичных троглобионтов (обитателей пещер) глаза могут совсем отсутствовать. Не имели глаз также представители вымершего палеозойского семейства Arthromygalidae из подотряда Liphistiomorphae (Petrunkevitch, 1923, 1953; Дубинин, 1962).
Глаза занимают передний участок головы - так называемую глазную область (или глазное поле) ее и располагаются здесь либо одной тесной группой, либо в 2-3 поперечных ряда. В последнем случае, если число глаз в переднем или заднем ряду равно 4, различают боковые (латеральные) и срединные (медиальные) глаза. Средний ряд, если имеется, всегда составляется только из 2 глаз. Вообще относительные размеры и расположение глаз очень сильно варьируют и характерны для каждого семейства пауков (рис. 70, 71, 75).
У многих пауков передние медиальные глаза темные, почти черные, а остальные - светлые, голубоватые. В таких случаях темные глаза обозначаются как "дневные", а светлые как "ночные"; это подразделение, однако, чисто условно и не связано с какими-либо функциональными особенностями тех и других глаз.
На самой передней, лицевой поверхности головы между глазами и хелицерами расположен наличник (clypeus). У большинства пауков наличник вертикальный (рис. 72), но у некоторых из них он принимает горизонтальное или почти горизонтальное положение. Ширина (или высота) наличника может быть различной и определяется расстоянием между передними глазами и основанием хелицер.
Основную часть вентральной стороны головогруди: занимает стернальный щит, или стернум (sternum), который образуется в результате слияния стернитов 4 сегментов ходильных ног (рис. 7). Тазики этих ног располагаются по краям стернума и соединяются с ним мягкой перепончатой мембраной. Задняя часть стернума у многих пауков имеет срединный заостренный вырост. Стернит педипальп обычно лежит свободно, не прирастая к стернуму, и образует нижнюю губу (labium), закрывающую вход в предротовую полость. Лишь у некоторых Mygalomorphae, а также у Filistatidae и Pholcidae из Araneomorphae нижняя губа полностью сливается со стернальным щитом.
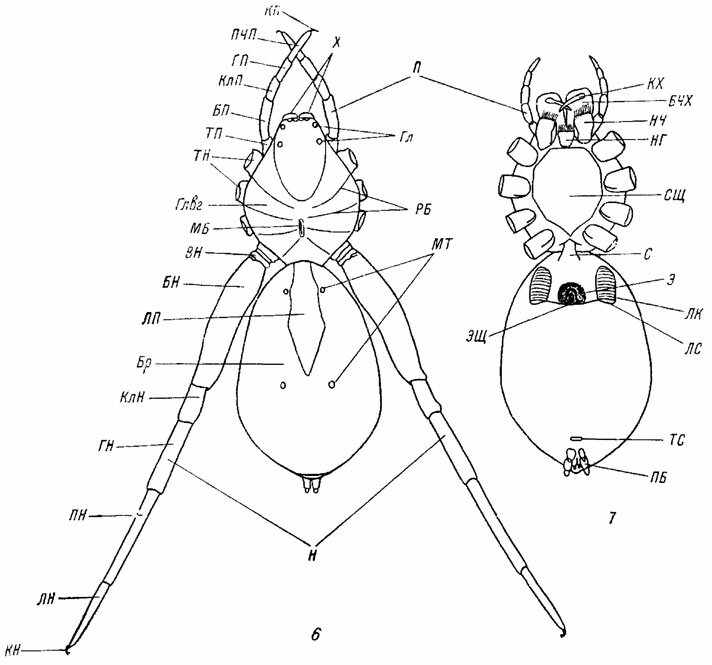
Рис. 6, 7. Схема строения тела паука Trockosa sp. Ориг. Рис. 6. Спинная сторона. Рис. 7. Брюшная сторона. БН - бедро ноги; БП - бедро пальпы; Бр - брюшко; БЧХ - базальный членик хелицер; ВН - вертлуг ноги; Гл - глаза; Глвг - головогрудь; ГН - голень ноги; ГП - голень пальпы; КлН - колено ноги; КлП - колено пальпы; КН - коготки ноги; КП - коготок пальпы; КХ - коготок хелицеры; ЛК - легочная крышечка; ЛН - лапка ноги; ЛП - ланцетовидное пятно; ЛС - легочная стигма; МБ - медиальная бороздка; МТ - мускульные точки; Н - ноги; НГ - нижняя губа; НЧ - нижние челюсти (максиллы); П - пальпы; ПБ - паутинные бородавки; ПН - пред-лапка ноги; ПЧП - последний членик пальпы; РБ - радиальные бороздки; С - стебелек; СЩ - стернальный щит; ТН - тазики ног; ТП - тазик пальпы; ТС - трахейная стигма; X - хелицеры; Э - эпигина; ЭЩ - эпигастральная щель
Предротовая полость спереди ограничивается хелицерами, а с боков прикрывается специальными лопастями тазиков педипальп, получившими название нижних челюстей, или максилл (maxillae)*. Развитие этих лопастей происходило уже после формирования отряда пауков, и поэтому наиболее примитивные группы (Liphistiomorphae и большинство Mygalomorphae) лишены их.
*(Термин "максиллы", который широко используется в систематике пауков, только условно может быть применен к челюстным лопастям педипальп, так как последние не имеют подвижного сочленения с тазиками и не принимают участия в раа-мельчении добычи (Иванов, 1965).)
Придатки головогруди. Головогрудь несет хелицеры, педипальпы и 4 пары ходильных ног.
Хелицеры (chelicerae) с помощью сочленовной мембраны подвижно сочленяются с головогрудью и состоят из двух члеников - базального членика и коготка (рис. 8- 10). Базальный, или основной, членик хелицер толстый и широкий. Внутри этого членика расположены мощные мышцы, приводящие в движение коготок, и ядовитая железа с ее протоком. У примитивных представителей отряда (у Lipkistiomorphae и большинства Mygalomorphae) ядовитая железа целиком располагается в базальном членике хелицер, но у остальных пауков большая часть ее лежит в головогруди (рис. 9, 10). Проток ядовитой железы открывается на конце изогнутого и заостренного коготка. Представители подотрядов Liphistiomorphae и Mygalomorphae имеют направленные вперед хелицеры с расположенными почти параллельно друг другу коготками (рис. 4, 45, 47). У пауков подотряда Araneomorphae хелицеры направлены вниз и более или менее перпендикулярны к продольной оси тела, а коготки хелицер расположены навстречу друг другу (рис. 7). В месте соединения коготка с базальным члеником имеется небольшой промежуточный склерит, который, возможно, представляет собой рудимент третьего промежуточного членика хелицер.
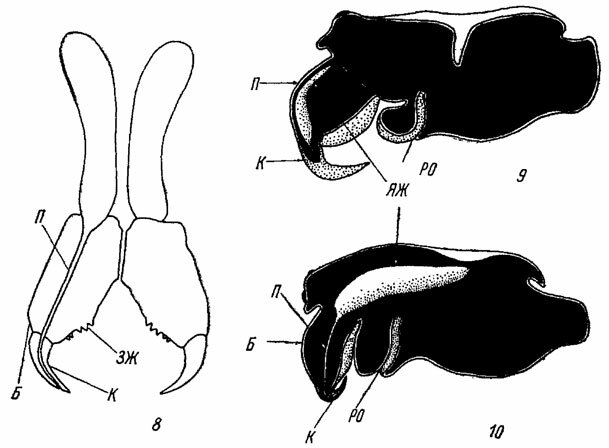
Рис. 8-10. Хелицеры и ядовитые железы пауков. По Мариковскому и Кроме, изменено. Рис. 8. Хелицеры тарантула Lycosa singoriensis Laxm. (Lycosidae). Рис. 9, 10. Схема расположения ядовитой железы: 9 - у Mygalomorphae; 10 - у Атапеоmorphae. В - базальный членик хелицеры; ЗЖ - зубцы желобка хелицеры; К - коготок; П - проток ядовитой железы; РО - ротовое отверстие; ЯЖ - ядовитая железа
В обычном состоянии коготок хелицер тесно прилегает к базальному членику, помещаясь в специальном желобке. По внутреннему (заднему) и наружному (переднему) краям этого желобка обычно расположены различные зубцы или крупные зубовидные выросты, форма и расположение которых имеют большое значение в систематике пауков.
Хелицеры в основном используются пауками для ловли, удерживания и умерщвления добычи. Через коготок секрет ядовитой железы вводится внутрь тела жертвы и вызывает ее гибель. У Ctenizidae хелицеры служат для выкапывания норок и снабжены на конце базального членика сильными копательными шипами (Wiehle, 1954). Тарантул (Lycosa singoriensis) при изготовлении норы также интенсивно работает хелицерами (Мариковский, 1956). Кроме того, хелицеры могут использоваться самками для перетаскивания яйцевых коконов (Pisaura, Pholcus, Micrommata) и самцами - для удерживания самки во время копуляции. В этом последнем случае хелицеры самцов сильно развиты, вооружены многочисленными зубцами и имеют очень длинные и толстые коготки (Tetragnathidae - рис. 605, Micryphanthidae - рис. 749).
Педипальпы (pedipalpi), или пальпы, значительно длиннее хелицер и по своему строению напоминают укороченные ходильные ноги (рис. 11). Они состоят из 6 члеников: тазика (соха), вертлуга (trochanter), бедра (femur), колена (patella), голени (tibia) и лапки (tarsus). Лишь у самок тропического семейства Symphytognathidae пальпы подвергаются сильной редукции и состоят только из одного членика. Тазики пальп у большинства пауков, как уже отмечалось, снабжены расширенными челюстными лопастями, которые по их внутреннему краю усажены многочисленными утолщенными волосками (рис. 11). Остальные членики пальпы покрыты простыми волосками, щетинками, а иногда даже толстыми шипами. Лапка обычно заканчивается 1-2 гребневидными коготками.
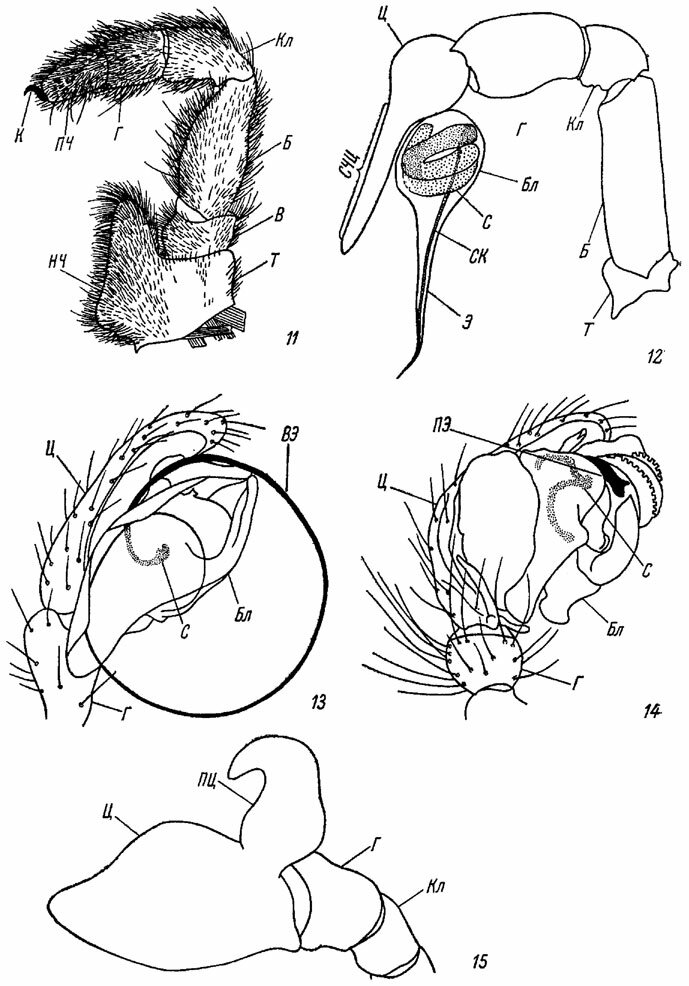
Рис. 11-15. Пальпы пауков. По Милло и Виле. Рис. 11. Пальпа самки Atypus muralis Bert. (Atypidae). Рис. 12. Пальпа половозрелого самца Scytodes lawrencei Less. (Sicariidae), Рис. 13-15. Последний членик пальпы половозрелого самца: 13 - Linyphia pusilla Sund. (Linyphiidae); 14 - кончик пальпы половозрелого самца Linyphia montana Cl.; 15 - то же Bathyphantes dorsalis Wid. (Linyphiidae), вид с дорсальной стороны. Б - бедро; Бл - бульбус; В - вертлуг; ВЭ - вводимый эмболюс; Г - голень; К - коготок; Кл - колено; НЧ - нижняя челюсть (максилла); ПЦ - парацимбиум; ПЧ - последний членик пальпы (лапка); ПЭ - присоединяемый эмболюс; С - спермофор; СК - семяизвергательный канал; СЧЦ - свободная часть цимбиума; Т - тазик; Ц - цимбиум; Э - эмболюс
Пальпы являются местом расположения различных рецепторов и в этом отношении аналогичны усикам насекомых. В ходьбе они не участвуют. У самцов всех пауков пальпы превращаются в сложно устроенные копулятивные органы.
Превращение пальпы в копулятивный аппарат сопровождается значительной модификацией концевых члеников и в особенности лапки, с помощью которой осуществляется перенос спермы в женское половое отверстие. У половозрелых самцов лапка (или последний членик пальпы) сильно расширяется и приобретает ложкообразную форму (рис. 13-16). Такая видоизмененная лапка пальпы получает название лодочки, или цимбиума (cymbium). У некоторых пауков наблюдается разрастание основания цимбиума и образование особого, иногда очень крупного придатка - парацимбиума (paracymbium, рис. 15, ПЦ).
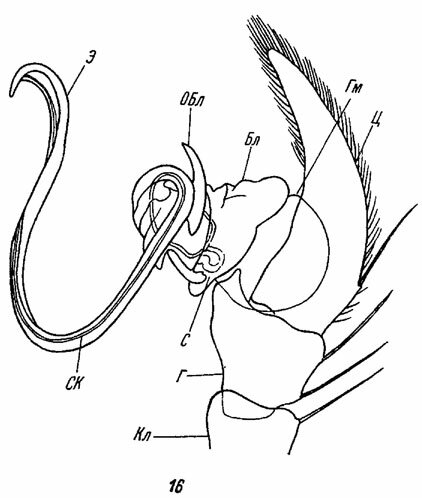
Рис. 16. Схема строения мужского копулятивного аппарата паука Agelena sp. По Петрункевичу. Бл - бульбус; Г - голень пальпы; Гм - гематодоха. Кл - колено; ОБл - отросток бульбуса; С - спермофор; СК - семяизвергательный канал; Д - цимбиум; 9 - эмболюс
На лодочке пальпы в специальной выемке ее (alveolus) располагается собственно копулятивный орган, проксимальный отдел которого представлен гематодохой (hematodocha), а дистальный - луковицей, или бульбусом (bulbus). Гематодоха - это тонкостенный мембранозный мешок, полость которого сообщается с полостью тела. При копуляции этот мешок заполняется гемолимфой и разворачивается вместе с бульбусом и его придатками. Бульбус сидит на вершине гематодохи и в покое располагается внутри выемки цимбиума. У Dysderidae, Sicariidae и некоторых других пауков, которые выделяются в особую группу семейств Haplogynae, бульбус устроен очень просто и представляет собой пузырчатый или грушевидный придаток последнего членика пальпы (рис. 12). У представителей семейств группы Entelegynae бульбус имеет более сложное строение и снабжен различными придатками, отростками и склеротизованными пластинками, которые служат для защиты и закрепления его на женском копулятивном аппарате (Bhatnagar a, Sadana, 1963). По мере эволюции пауков даже в пределах каждого семейства отмечается параллельное усложнение строения копулятивных органов самцов и самок (Вагнер, 1886в; Levi, 1961; Merrett, 1963).
Бульбус последнего членика пальпы самцов заканчивается специальным стилетом, или эмболюсом (emboli), который выполняет функцию пениса (рис. 16). Существует два основных типа строения эмболюса: вводимый и присоединяемый эмболюс (Wiehle, 1961). Вводимый эмболюс очень длинный и часто закрученный в виде спирали (рис. 13); при копуляции он вталкивается в половое отверстие самки. Присоединяемый эмболюс обычно короткий и снабжен многочисленными железами, которые облегчают его закрепление на копулятивном аппарате самки (рис. 14).
Внутри бульбуса расположен спермофор, который продолжен в семяизвергательный канал, проходящий внутри эмболюса и открывающийся наружу на его кончике. Перед копуляцией самец паука изготовляет из паутины небольшую треугольную сперматическую сеточку и с ее помощью заполняет спермофор семенной жидкостью (см. стр. 18). В момент копуляции сперма, заполняющая спермофор, проходит через семяизвергательный канал эмболюса в семеприемники самки. После копуляции кончик эмболюса часто обламывается и остается в каналах семеприемников самки (Biasi, 1962; Abalas a. Baez, 1963; Wiehle, 1967).
Голень и остальные членики пальп самцов сохраняют свое первоначальное строение, но часто имеют специфические отростки, шипы или зубцы, которые широко используются в систематике пауков.
Ходильные ноги присутствуют всегда в числе 4 пар, которые обозначаются римскими цифрами (ноги I, II, III и IV) и имеют однотипное строение у всех пауков. Относительная длина каждой ноги описывается так называемой формулой ног, в которой порядковые номера конечностей расположены по степени уменьшения их величины. Так, например, формула 2.1.4.3 означает, что вторая нога - наиболее длинная, первая - длиннее четвертой и третьей, а третья - наиболее короткая.
Каждая нога паука состоит из 7 члеников: тазика, вертлуга, бедра, колена, голени, предлапки (metatarsus) и лапки (рис. 17). У Mygalomorphae и у большинства пауков-скакунчиков (сем. Salticidae) ноги относительно короткие и толстые. У Pholcidae все членики ног, наоборот, тонкие и длинные, а лапка имеет большое число вторичных, или ложных, члеников.
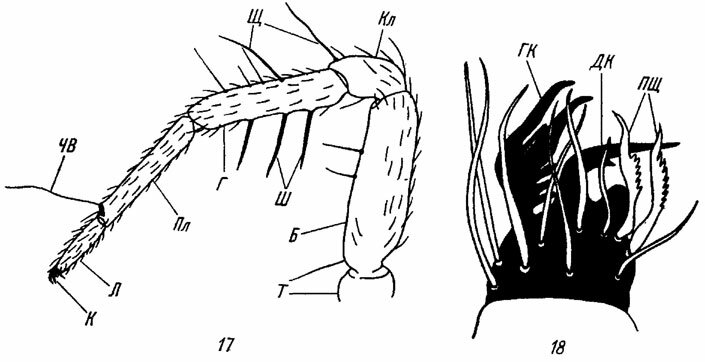
Рис. 17, 18. Строение ноги пауков. По Виле и Мариковскому. Рис. 17. Передняя нога Minicia marginella Wid. (Micryphantidae). Рис. 18. Кончик лапки последней ноги каракурта Latrodectus tredecimguttatus Rossi (Theridiidae). Б - бедро; Г - голень; ГК - главный коготок; ДК - добавочный коготок; К - коготки лапки; Кл - колено; Л - лапка; Пл - предлапка; ПЩ - прядильные щетинки; Т - тазик; ЧВ - чувствительный волосок; Ш - шипы; Щ - щетинки
Концевая часть лапки (praetarsus) несет два главных коготка, снабженных зубчиками. Между этими парными коготками расположен особый выступающий придаток, называемый эмподием (empodium). У многих пауков эмподий превращен в маленький дополнительный коготок, лишенный зубцов (рис. 18). В подотряде Araneomorphae появление допблнительного коготка характерно для семейств, объединяемых в одну естественную группу Trionycha. У пауков-тенетников, которые в большинстве своем относятся к этой группе, иногда наблюдается превращение терминальных шипов задних лапок в когтеобразные, S-образные или пильчатые структуры (рис. 18), приспособленные для передвижения паука по паутине или для изготовления паутинных нитей ("прядильные щетинки").
Ноги пауков покрыты волосками, особенно многочисленными у обитающего в воде паука-серебрянки (Argyroneta aquatica Cl.). По своему строению эти волоски могут быть простыми, перистыми или чешуйчатыми. Волоски вентральной стороны предлапки и лапки иногда утолщены и расположены в виде плотной щеточки (скопула - scopula, рис. 61). Волоски, образующие скопулу, по-видимому, способны выделять клейкий секрет, и поэтому пауки, обладающие ими (некоторые Thomisidae, Ctenidae, Dipluridae и др.), могут ползать по гладкой вертикальной поверхности (Нотапп, 1957).
В группе семейств Cribellatae на дорсальной стороне предлапки IV развивается 1-2 продольных ряда специфических прядильных волосков, формирующих каламиструм (рис. 54, 55). Образование каламиструма связано с видоизменением паутинного аппарата (появление крибеллума - см. стр. 14) и выпряданием особых "крибеллятных" паутинных нитей.
Многие членики ног вооружены утолщенными волосками - щетинками и подвижными сочлененными у основания шипами. Кроме того, ноги пауков снабжены длинными и очень тонкими чувствительными волосками - трихоботриями (рис. 17, ЧВ). Расположение, относительные размеры и форма щетинок, шипов или чувствительных волосков имеют очень большое значение в родовой систематике пауков (см., например, семейства Micryphantidae, Linyphiidae, Lycosidae и др.).
В ходьбе и беге пауков участвуют, как правило, поочередно все 4 пары ходильных ног. Обычно передние ноги служат для подтягивания тела паука, а задние - для подталкивания его. Однако у Рalpimanidae и Salticidae передняя пара ног не принимает участия в передвижении паука: она приподнимается над субстратом и вытягивается вперед наподобие пальпы. Некоторые пауки могут прыгать, отталкиваясь от почвы задними ногами.
Стебелек (petiolus, рис. 7), соединяющий головогрудь с брюшком, представляет собой тонкий и длинный первый сегмент опистосомы. У пауков, имитирующих муравьев, этот сегмент особенно сильно удлиняется и хорошо заметен при рассматривании паука сверху (рис. 5). У большинства остальных пауков дорсальная часть стебелька прикрывается брюшком и сверху незаметна.
Брюшко (abdomen) у подавляющего большинства пауков не имеет первичной наружной сегментации и представляет собой цельный мешок, ограниченный мягкой, способной растягиваться кутикулой. Только у всех ископаемых палеозойских пауков и у современных представителей подотряда Liphistiomorphae брюшко сохраняет наружную сегментацию в виде 11 склеротизованных тергитов и отвечающих им более мягких стернитов (рис. 4). Обширные, сильно разросшиеся плейриты брюшных сегментов у Liphistius занимают боковые поверхности брюшка. В подотрядах Mygalomorphae и Araneomorphae членистое строение брюшка наблюдается лишь у только что вылупившихся из яйца паучков (Millot, 1931; Petrunkevitch, 1933; Holm, 1940). В более старших возрастах, когда наружная сегментация брюшка исчезает, следы ее могут сохраняться в виде метамерного расположения дорсальных волосков и темных пятен рисунка (Crome, 1955а, 1955b).
Подробный анализ сегментации тела пауков позволил установить, что как у Liphistiomorphae, так и у Mygalomorphae и Araneomorphae в состав брюшка входят 6 сегментов мезосомы и 5 сегментов метасомы, т. е. всего 11 опистосоматических сегментов, однако у высших пауков вентральные части 6 задних сегментов редуцируются (Kastner, 1937; Millot, 1949; Иванов, 1965).
Среди пауков нередки случаи образования твердых вторичных склеритов на мягкой поверхности брюшка. Подобные склериты имеются, например, у некоторых Oonopidae, Micryphantidae и Gnaphosidae (рис. 783). Особенно часто наблюдается формирование дорсального склерита то в виде небольшой треугольной пластинки, то в виде обширного щита. Реже развивается также вентральный склерит. Крупные щиты этих склеритов, получившие соответственно названия дорсального и вентрального скутумов, образуют твердый панцирь, защищающий брюшко от механических повреждений.
Дорсальная поверхность брюшка имеет очень простое строение. У многих пауков, обладающих светлой окраской, на дорсальной поверхности брюшка располагается темное продольное ланцетовидное пятно, которое создается просвечивающей через покровы сердечной трубкой (рис. 6, ЛП). Разбросанные повсюду маленькие кутикулярные впячивания - мускульные точки брюшка отвечают местам прикрепления дорсовентральных мышц. Брюшко оканчивается маленьким анальным бугорком.
Вентральная поверхность брюшка имеет гораздо более сложное строение, так как здесь располагаются половое отверстие, копулятивный аппарат самки, стигмы и паутинные бородавки.
Половое отверстие самок и самцов пауков находится у Liphistimorphae и Araneomorphae в специальной эпигастральной борозде, расположенной у основания брюшка и отвечающей границе II и III опистосоматических сегментов (рис. 7). Только у Те-tragnathidae половое отверстие вторично сдвинуто назад за пределы борозды, а у Му-galomorphae и у Dysderidae из Araneomorphae эпигастральная борозда вообще отсутствует.
У самок большинства пауков неподалеку от полового отверстия находятся самостоятельные парные наружные отверстия семеприемников, через которые при копуляции сперма проходит из эмболюса самца в семеприемники (receptacula seminis) самки или их каналы (рис. 20-22). Здесь, в семеприемниках самки, сперма может храниться длительное время, выходя по специальным оплодотворительным каналам в момент откладки яиц. По мнению П. И. Мариковского (1956), выход спермы в оплодотворительный канал во время яйцекладки обусловливается проникновением гемолимфы в семеприемник через специальные многочисленные поры на дистальном конце его.
У Mygalomorphae, а также у Filistatidae, Sicariidae, Dysderidae и Tetragnathidae из Araneomorphae отверстия семеприемников открываются непосредственно на вентральной поверхности брюшка и не имеют каких-либо дополнительных структур, выполняющих функцию совокупительных органов. У всех остальных пауков эти отверстия связаны с наружной склеротизованной площадкой эпигины (epigyne), расположенной над эпигастральной щелью (рис. 19). В типичном случае основная часть эпигины прикрывается медиальной пластинкой, имеющей различное строение и прикрывающей частично или полностью ямки эпигины (рис. 19). Однако иногда медиальная пластинка может отсутствовать, и тогда одна или несколько ямок эпигины открыты полностью.
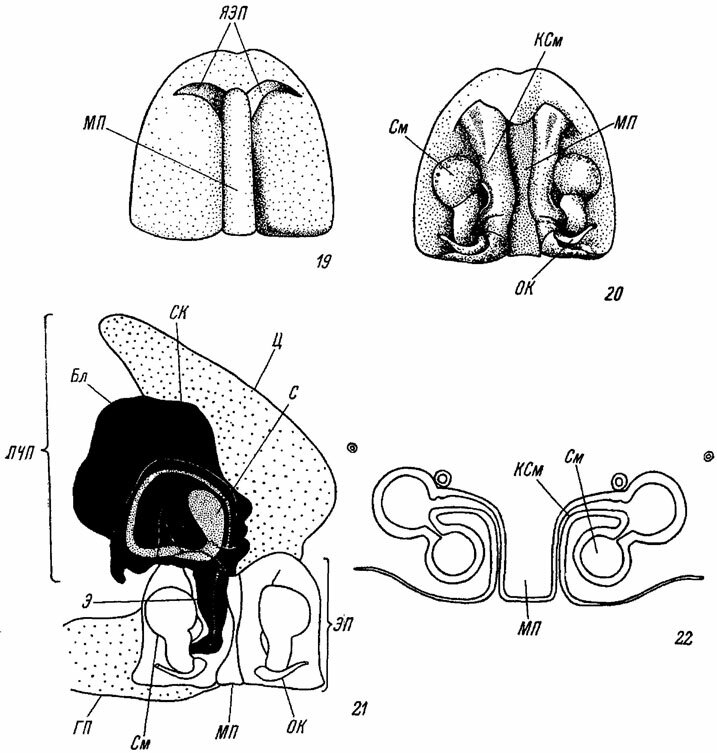
Рис. 19-22. Схема строения эпигины и семеприемников у самки Cupiennius salei Keys. (Ctenidae). По Мелхерсу. Рис. 19, 20. Эпигина: 19 - вид снаружи; 20 - вид изнутри. Рис. 21. Положение копулятивного органа самца на эпигине самки во время копуляции. Рис. 22. Поперечный срез через эпигину. Бл - бульбус; ГП - голень пальпы; КСм - канал семеприемника; МП - медиальная пластинка; OК - оплодотворительный канал; ПЧП - последний членик пальпы; С - спермофор; СК - семяизвергательный канал; См - семеприемник; Ц - цимбиум;. Э - эмболюс; ЭП - эпигина; ЯЭП - ямка эпигины
Усложнение строения эпигины наблюдается особенно часто у Linyphiidae, Aranedae и у ряда других семейств высших пауков. Первоначально наблюдается образование скапуса (scapus) - специального хитинового возвышения, разрастающегося спереди назад и прикрывающего сверху всю эпигину с ее медиальной пластинкой и ямками (рис. 721-726), На заднем крае скапус может переходить в длинный узкий отросток, достигающий иногда середины брюшка (рис. 642). С другой стороны непосредственно на поверхности скапуса может, наблюдаться образование структуры III порядка в виде более или менее длинного и гибкого придатка - клавуса (clavus), частично закрывающего скапус (рис. 588, 592, 595). В некоторых случаях наблюдается также разрастание заднего края эпигины в виде отростка, получившего название пармулы (parmula). Этот отросток заднего края эпигины иногда может далеко заходить за эпигастральную щель (рис. 722, 723).
Все описанные выше разнообразные усложнения строения эпигины имеют несомненно адаптивное значение и сопровождаются соответствующими изменениями в структуре копулятивного органа самцов. Морфологические особенности эпигины (наличие отростков, форма медиальной пластинки, расположение ямок) обеспечивают тонкую и точную подгонку ее к сложному копулятивному аппарату самца наподобие системы замка и ключа (Bnatnagar a. Sadana, 1963). В момент копуляции эмболюс самца вводится в семеприемник самки через одно из отверстий каналов семеприемника, а бульбус своими отростками закрепляется за соответствующие впадины (=ямки) и выросты эпигины самки (рис. 21).
Стигмы (stigmata) - дыхательные отверстия .расположены на II и III сегментах опистосомы и связаны либо с легкими, либо с трахеями. Легочные стигмы обычно широкие, щелевидные, а трахейные стигмы - узкие, малозаметные. Легкие всегда помещаются в передней половине брюшка и легко опознаются по так называемым легочным крышечкам, которые резко отличаются от окружающей кутикулы темной окраской, наличием поперечных морщинок и отсутствием волосков (рис. 7). Трахеи, как правило, располагаются ближе к концу брюшка.
Обширную группу составляют четырехлегочные пауки (Tetrapneumones), у которых трахеи отсутствуют, а органами дыхания служат две пары легких, открывающихся двумя парами легочных стигм. К ним принадлежат Liphistiomorphae, Mygalomorphae - и сем. Hypochilidae из Araneomorphae. Передняя пара стигм в этом случае располагается на уровне полового отверстия, а задняя пара - несколько отступя к середине брюшка.
У двулегочных пауков (Dipneumones) вторая пара легких замещается трахеями (большинство Araneomorphae) или полностью редуцируется (Pholcidae, Diguetia, Sicarius). Некоторые представители этой группы (Oonopidae и Dysderidae) имеют подобные легочным стигмам парные трахейные стигмы, расположенные вблизи эпигастральной борозды (рис. 48). У остальных двулегочных пауков в результате слияния парных стигм образуется непарная трахейная стигма, которая обычно смещается далеко-назад и располагается непосредственно впереди паутинных бородавок (рис. 7).
Особую группу безлегочных пауков (Apneumones) составляют пауки трех небольших тропических семейств (Caponiidae, Telemidae и Symphytognathidae), у которых первая пара легких также замещена трахеями. Трахейные стигмы у этих пауков всегда парные.
Паутинные бородавки (mamillae) представляют собой сильно видоизмененные ножки IV и V сегментов опистосомы и располагаются на вентральной стороне брюшка, в большинстве случаев - на самом конце его (рис. 7). Только у Liphistius все паутинные бородавки смещены к середине брюшка (рис. 4), а у Brachybothrium (Ctenizidae), Myandra (Clubionidae) и Zimiris (Prodidomidae) здесь помещается передняя пара бородавок.
Четыре пары, т. е. полный набор паутинных бородавок, среди современных пауков обнаруживаются лишь у Liphistiomorpha (рис. 4, 23, А). Представители рода Heptathele из Araneomorpha имеют 7 паутинных бородавок (рис. 23, Б), а все остальные пауки - 6 и менее. Очевидно, по мере эволюции в разных группах пауков постепенно происходила параллельная олигомеризация - уменьшение числа паутинных бородавок (рис. 23). Крайняя степень олигомеризации достигается, с одной стороны, у Diplothele,. Anisaspis и Anisaspoides среди Mygalomorphae (рис. 23, В-Д), а, с другой стороны, у Palpimanidae и у некоторых Zodariidae среди Araneomorphae (рис. 23, К). У этих пауков остается всего лишь одна пара бородавок. Полной редукции всех паутинных бородавок, однако, никогда не наблюдается.
Почти все пауки европейской фауны имеют 3 пары паутинных бородавок (рис. 24).. В некоторых случаях сохраняются также гомологи исчезнувшей пары бородавок - колюлус и крибеллум. Передние паутинные бородавки обычно крупные, почти всегда двучлениковые и широко раздвинутые. Среди тропических видов у большинства Mygalomorphae и у некоторых примитивных форм Araneomorphae эти бородавки отсутствуют. Средние (медиальные) паутинные бородавки очень мелкие, одночлениковые, тесно сближенные. Наконец, задние паутинные бородавки - крупные 2-, 3- или даже 4-члениковые. Обычно паутинные бородавки располагаются одной компактной группой, причем медиальные бородавки прикрываются двумя остальными парами и плохо заметны. Только у Hahnia все три пары паутинных бородавок вытянуты в один поперечный ряд (рис. 444).
Между основаниями передних паутинных бородавок у многих пауков лежит маленький придаток - колюлус (colulus) (рис. 24) - рудимент дополнительной передней пары медиальных бородавок Liphistius (Thorell, 1869). Небольшой ситовидный склерит - крибеллум (cribelhim, рис. 51), свойственный паукам группы Cribellatae, также представляет собой производное этих исчезнувших бородавок и подобно последним располагается перед остальными бородавками. Крибеллум часто имеет парную природу и подразделяется на два обособленных отдела (Ergatis, Amaurobius, рис. 52).
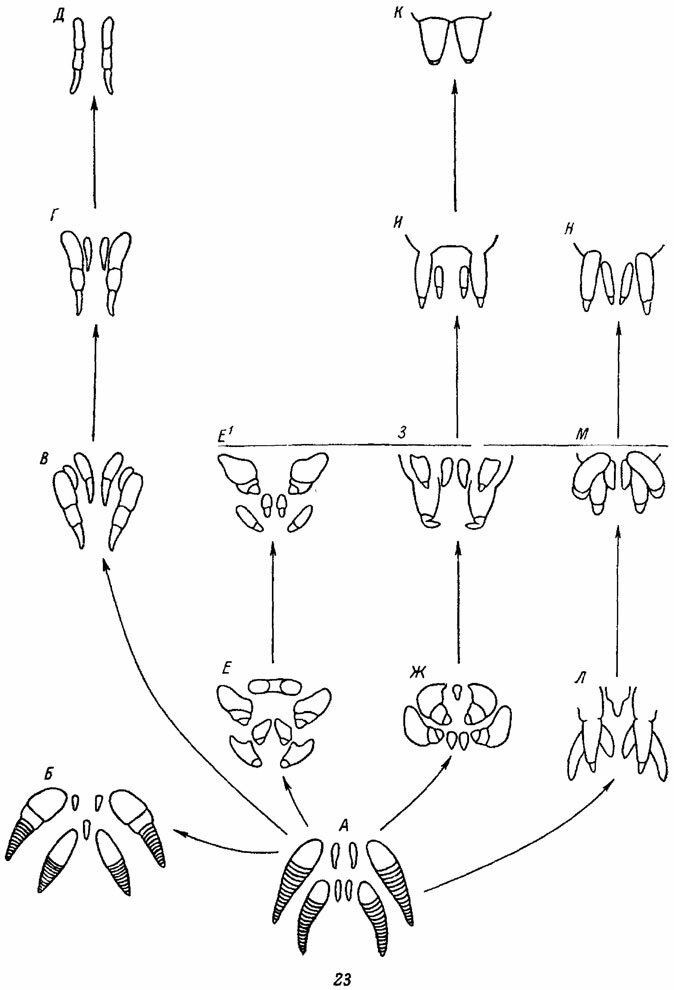
Рис. 23. Параллельная олигомеризация паутинных бородавок в разных группах пауков. По Иванову. А - Liphistius; В - Heptathele; В - Atypus; Г - большинство Mygalomorphae; Д - Barychelidae, Е - Stegodyphus, самка; Е1 - то же, самец Ж - Araneus; З - Uroctea; И - некоторые Entelfgynae; К - Zodariidae; Л - Loxosceles; М - многие Haplogynae; H - Hadrotarsus
На паутинных бородавках расположены паутинные трубочки (fusulae), через которые выходят нити секрета желез, застывающие на воздухе в виде паутины. Расположенные в брюшке паутинные железы пауков очень разнообразны и многочисленны. Так, самка паука-крестовика (Araneus diadematus Сl.) имеет в общей сложности 480-560 мелких и около 20 крупных паутинных желез, относящихся к 6 основным типам (Apstein, 1889; Hopfmann, 1935; Dumitresco, 1941). Недавно был описан еще один, седьмой, тип паутинных желез Araneidae (Sekiguchi, 1952). Паукам группы Cribellatae свойственны особые паутинные железы, которые открываются на крибеллуме, также снабженном паутинными трубочками. Общее число трубочек и паутинных желез крибеллума обычно очень велико, и, например, у самок Stegodyphus lineatus Latr. достигает 9600.
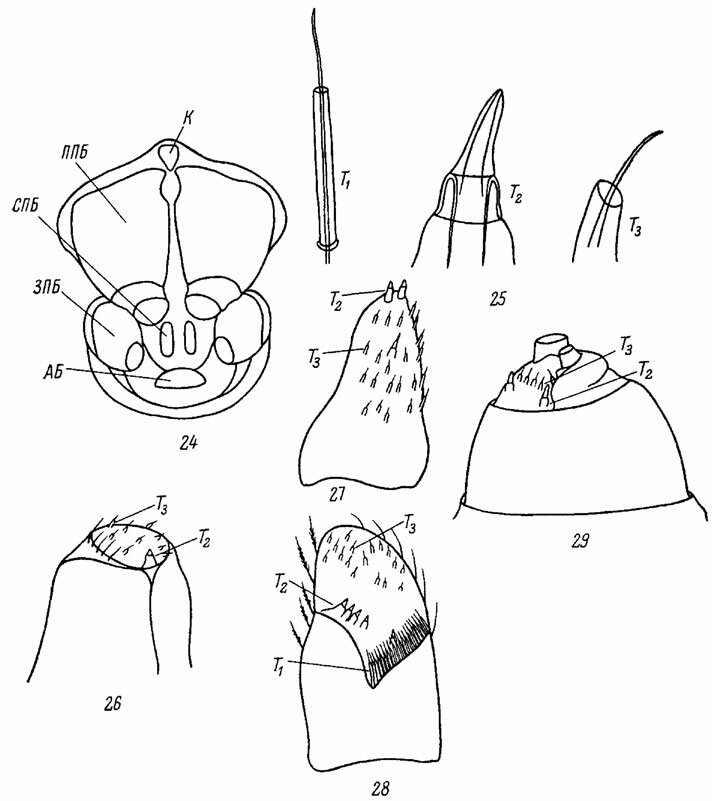
Рис. 24-29, Паутинные бородавки пауков. По Симону, Виле и ориг. Рис. 24. Паутинные бородавки Araneus sp. Рис. 25. Паутинные трубочки Araneus sp. Рис. 26-28. Схема расположения паутинных трубочек на паутинных бородавках Araneus sp.: 26 - на передней; 27 - на средней; 28 - на задней. Рис. 29. Задняя паутинная бородавка Steatoda sp. АБ - анальный бугорок; ЗПБ - задние паутинные бородавки; К - колюлус; ППБ - передние паутинные бородавки; СПБ - средние паутинные бородавки; Т1, Т2, Т3 - различные типы паутинных трубочек
Каждая паутинная трубочка связана только с одной железой и состоит из двух частей: более толстой базальной и более тонкой концевой. У Araneus описаны три основных типа паутинных трубочек (Simon, 1892). Базальная часть трубочек 1-го типа представлена высоким- узким цилиндриком с абсолютно параллельными краями; концевая часть трубочки тонкая и прямая (рис. 25, Т1). Трубочки 2-го типа ("паутинные конусы") очень массивные даже в своей концевой части и сильно склеротизованные у основания (рис. 25, Т2). 3-й тип трубочек имеет небольшую конусовидную базальную часть и тонкую изогнутую концевую часть (рис. 25, Т3).
Каждая паутинная бородавка характеризуется определенным числом трубочек того или иного сорта (McCook, 1889-1892). Так, например, передние паутинные бородавки Araneus имеют 60-70 трубочек 2-го типа и только одну крупную трубочку 3-го типа (рис. 26). Средние паутинные бородавки этих же пауков также не обладают трубочками 1-го типа (рис. 27), и лишь задние бородавки имеют полный набор трубочек всех трех типов (рис. 28). Форма и число паутинных трубочек часто различны в разных семействах пауков. У Araneidae трубочки малы и многочисленны, а, например, у Gnaphosidae, напротив, представлены в небольшом числе и сравнительно крупны (Иванов, 1965), У Tegenaria (Agelenidae) трубочки всех трех паутинных бородавок подобны и, вероятно, принадлежат к одному и тому же типу. Наоборот, у Theridiidae кроме трех описанных выше типов трубочек имеются еще очень крупные "паутинные цилиндры", лишенные концевой части (Hopfmann, 1935; Wiehle, 1949).
Брюшко заканчивается небольшим дву- или трехчленистым анальным бугорком, у основания которого лежит анальное отверстие (рис. 7). В некоторых случаях, например у Oecobiidae, анальный бугорок снабжен густым венчиком очень длинных волосков (рис. 53).
Звуковые (стридуляционные) органы у пауков разных семейств расположены на разных частях тела и состоят из зубчиков или шипов и из противопоставленных им плотных волнистых или сморщенных поверхностей. Звуки возникают при движении одной части органа относительно другой. У Mygalomorphae звуковые органы обычно представлены видоизмененными щетинками, расположенными на хелицерах и на бедренных члениках пальп. Трение этих щетинок друг о друга производит ясно слышимые звуки. У представителей Araneomorphae встречаются самые разнообразные типы строения звуковых органов. Наиболее известны звуковые аппараты самцов многих Theridiidae, расположенные на соприкасающихся поверхностях брюшка и головогруди. В этом случае передняя сторона брюшка, над стебельком, несет ряд зубчиков, а головогрудь - несколько очень тонких бороздок (рис. 361). Звуки, которые производит аппарат этого типа, очень слабы и не воспринимаются человеком. У Antistea elegans Black. (Agelenidae) звуковой аппарат состоит из мелких рассеянных зубчиков на задней части головогруди и двух заметных пятен из плотно прижатых расширенных щетинок на передней поверхности брюшка (Helsdingen, van, 1963). У самцов Lepthyphantes (Linyphitdae), а также у Sicariidae части звукового аппарата расположены с одной стороны на хелицерах, а с другой стороны - на бедрах пальп. Наконец, у самцов Rhaebothorax brocchus L. Koch и Eboria caliginosa Falc. (Micryphantidae) трущаяся поверхность звукового органа представлена легочными крышечками, которым противопоставлен зубовидный отросток тазиков последней пары ног.
|
ПОИСК:
|
 На сайте https://cheremhovosm.date большой выбор девушек.
На сайте https://cheremhovosm.date большой выбор девушек. При использовании материалов сайта активная ссылка обязательна:
http://paukoobraznye.ru/ 'Паукообразные: клещи, пауки, скорпионы'