Биология
Поведение пауков отличается необычайной сложностью. В период размножения, при постройке логовищ и сетей, при ловле добычи пауками можно наблюдать такие интересные явления, как замечательные "свадебные танцы", сопровождающиеся иногда поднесением "свадебных подарков", своеобразный способ копуляции, связанный с использованием пальп, функционирующих в качестве копулятивных органов самца, высокоразвитые строительные инстинкты и инстинкты заботы о потомстве, многообразные способы охоты и т. д. Изучение этих явлений, представляющих в биологическом отношении совершенно исключительный интерес, было начато еще в прошлом столетии и успешно продолжается в настоящее время. Полученные в этом направлении результаты сведены в обширных монографиях Комстока (Comstock, 1912), Сэвори (Savory, 1928), Нильсена (Nielsen, 1928-1932), Бристоу (Bristowe, 1939-1941), Тильквина (Tilquin, 1942), П. И. Мариковского (1956), А. В. Иванова (1965) и в ряде журнальных статей (см. библиографический указатель Бонне (Bonnet, 1945-1961) и список литературы на стр. 38).
Биология размножения. В зависимости от длительности периода половой зрелости, копуляции и яйцекладки все пауки могут быть разбиты на три основные группы. Эврихронные виды спариваются и откладывают яйца в течение длительного периода - всего лета и осени (Pachygnatha Clercki Sund.), осени и всей зимы (Leptyphantes cristatus Menge) или, в редких случаях, в течение всего года (Porrhomma egeria Sim.). Время половозрелости у стенохронных видов ограничено 1-2 месяцами или даже несколькими неделями в году. Так, в ГДР Uloborus walckenaerius Latr. размножается только в мае, Diaea dorsal a Fabr. - в мае и июне, a Xysticus sabulosus С. L. Koch - в октябре. У диплохрондых видов наблюдаются два периода размножения, которые приходятся на весну и осень (Coelotes inermis L. Koch) или на лето и зиму (Micrargus herbigradus Blackw.). Возможно, в особую группу следует выделить сравнительно немногие виды пауков с эврихронными самками и стенохронными самцами, К числу таких видов относится, например, Pirata latitans Blackw.
Перед спариванием самцы пауков заполняют спермой бульбус совокупительного аппарата, используя для этого особую сперматическую сеточку. Только у Segestria самец наполняет спермофор непосредственно из полового отверстия (Gerhardt и. Kastner, 1937-1938; Klein, 1953). Сперматическая сеточка бывает треугольной (Theridium, Araneus, Linyphia), четырехугольной (Его, Tegenaria) или даже линейной (Scytodes, Palpimanus). Самец выпускает на сеточку капельку семенной жидкости и погружает в нее концы пальп. Сперма заходит в спермрфоры бульбуса через особый канал, который впервые был описан П. И. Мариковским (1956).
Способ заполнения спермой копулятивного аппарата и положение самца во время этой операции характерны для каждого семейства или даже для каждого рода пауков. Так, например, самец каракурта (Latrodectus tredecimguttatus Rossi) выплетает на паутине самки П-образную нить с несколькими перемычками и удерживает ее в третьей паре ног. Затем самец выпускает капельку семенной жидкости на поперечную перекладину П-образной паутинки, которая после этого переносится вперед на уровень пальп. Последние поочередно приставляются к капельке и заполняются семенем (Мариковский, 1956). Подобным же образом поступают пауки Oonops и Scytodes, однако у них самец пользуется не П-образной паутинкой, а одной-единственной нитью (Gerhardt, 1928). С другой стороны, пауки сем. Agelenidae, например, делают плотную сперматическую сеточку и заполняют бульбусы просачивающимся сквозь нее семенем.
Особенно сложную сперматическую сеточку делают некоторые представители сем, Aviculariidae (подотряд Mygalomorphae), которые, чтобы предотвратить вытекание семени из сеточки, изготовляют "крепление" из паутины, выделяемой специальными железами "вентрального паутинного поля", расположенного между передними легкими (Melchers, 1964).
Еще более разнообразны способы "ухаживания" самцов, которые непосредственно предшествуют копуляции и, по-видимому, направлены на преодоление хищнических инстинктов самки (Сычевская, 1935). Движения "ухаживания", или "брачные танцы", пауков очень различны, но почти всегда они сводятся к вибрациям пальп и ног и к ритмическим подергиваниям брюшка. Самцы некоторых пауков (Lycosidae, Oxyopidae и Salticidae), обладающие пестрой окраской и особыми "украшениями" из ярких, волосков, принимают перед самкой своеобразные позы и демонстрируют свои замечательные "украшения". Самцы пауков-тенетников иногда делают по соседству с тенетами самки маленькие брачные сети, на которые они пытаются заманить самку своими "танцами". Наконец, самцы, имеющие звуковые органы, пользуются ими во Время "ухаживания".
Поднесение самцами самкам "свадебных подарков" в виде опутанной паутиной добычи наблюдается у Meta segmentata С1. и у Pisaura mirabilis Cl. У последнего вида самец перед спариванием ловит и умерщвляет муху, плотно запутывает в паутину и преподносит самке. Пока самка высасывает муху, самец успевает спариться с ней (Спасский, 1935; Schmidt, 1955).
Собственно спаривание пауков сводится к введению эмболюса пальпы самца в половые пути самки. При этом через отверстие на вершине эмболюса семенная жидкость проникает в семеприемник самки и хранится там вплоть до момента оплодотворения яиц. Обычно при, спаривании самец пользуется поочередно одной из пальп и лишь в редких случаях (у большинства Dysderidae и у некоторых Sicariidae, Oonopidae, Pholcidae) вводит оба эмболюса одновременно. Позы копулирующих партнеров весьма разнообразны и характерны для каждого семейства. Иногда положение полов при копуляции различно даже у близких видов. Так, например, самец Araneus pallidus Ol. обхватывает ногами головогрудь самки и вводит эмболюс в эпигину снизу, а у Л. diadematus Cl., самец обхватывает брюшко самки и вводит эмболюс в эпигину сверху (Grasshoff, 1964).
Во время спаривания самка часто впадает в состояние каталепсии, которое иногда бывает очень глубоким. После копуляции она выходит из этого состояния и во многих случаях высасывает самца (у Argiope bruennichi Scop., Cyrthophora, Theridium, Latrodectus). Однако у целого ряда видов (Araneus, Lycosa и др.) самец успевает убежать и не всегда делается жертвой самки. Самки Tegenaria вообще не стремятся убивать самцов при их приближении, но продолжительные поиски самок, "ухаживание" и копуляция настолько изнуряют самцов, что они обычно погибают во время спаривания или сразу после него (Savory, 1961). У пауков-скакунчиков (сем. Salticidae) самка также не проявляет агрессивности по отношению к самцу.
Яйцекладка происходит обычно спустя 2-6 недель, а у Atypus даже спустя 8-9 месяцев после спаривания (Potzsch, 1963). Яйца откладываются внутрь кокона, который изготовляется из паутины, выделяемой специальными трубковидными железами самок. У большинства пауков в коконе содержится несколько десятков яиц., но у Argiope bruennichi Scop, и у Theridium tepidariorum С. L. Koch их несколько сотен (Wiehle, 1931а, 1937). Количество яиц в коконах тарантула (Lycosa singoriensis Laxra.) и каракурта (Latrodectus tredecimguttatus Rossi) также колеблется от 300 до 700 (Мариковский, 1956). Однако некоторые типы коконов имеют незначительное число яиц: только 2 яйца у Oonops (Savory, 1928), 3 - у Pholcomma (Holm, 1940) и 8-9 - у Archaea (Millot et Legendre, 1964).
Простейший кокон Pholcus и Heliophanus состоит из немногих паутинных нитей, едва скрепляющих яйца. У остальных пауков кокон имеет достаточно прочные, иногда двойные стенки из паутины. При постройке такого кокона самка изготовляет первоначально основную пластинку, снабженную рыхлым краевым валиком, и откладывает на нее яйца. Затем основная пластинка с яйцами прикрывается сверху плотной кроющей пластинкой (рис. 35).
У Agelenidae, Dysderidae, Gnaphosidae и у многих других пауков готовый кокон имеет уплощенную или чечевицеобразную форму и плотно прикрепляется к субстрату (рис. 31, 34). Если при этом кокон располагается открыто, то его наружная оболочка часто маскируется кусочками почвы, мелкими веточками или песчинками. У Lycosidae изготовленный кокон отдирается от субстрата и поверх обеих пластинок оплетается паутинными нитями, приобретая шарообразную или слегка сплющенную форму (рис. 65). Подобный шарообразный кокон, свойственный кроме Lycosidae также Spa-rassidae, Ctenidae, Araneidae и другим паукам, в некоторых случаях снабжается на вершине заостренным паутинным бугорком и превращается в грушевидный кокон (рис. 30).
В ряде случаев строение кокона усложняется за счет появления вторичных оболочек. Так, например, у Argiope кокон состоит из двух плотных оболочек с толстым слоем рыхлой хлопковидной ткани между ними (Crome u. Сгоше, 1961). Если между оболочками такого двухслойного кокона остается свободное пространство, получается многокамерный кокон, обнаруженный, например, у видов рода Philodromus (рис. 36). У Micaria (Clubionidae) вторичная, наружная оболочка кокона образуется за счет паутинной стенки "гнезда", в котором у других пауков помещается самка вместе с коконом. В результате возникает гнездо-кокон, разделенное на две части: в одной половине его сидит самка, а другая половина занята коконом с яйцами (рис. 37). Подобное же происхождение имеет "сложный кокон" Agroeca brunnea Black, из того же семейства, но полость гнезда-кокона здесь нежилая (рис. 33, 38).
Сравнительно немногие виды пауков не проявляют инстинкта заботы о потомстве и не охраняют свои коконы. Таковы виды рода Zelotes, у которых коконы имеют плотные стенки и надежно замаскированы посторонними частицами (Nielsen, 1928-1932). Однако большинство пауков не только охраняют свои коконы, помещая их в тенетах или в специальных гнездах, но и ухаживают за ними. В зависимости от температуры воздуха и степени нагрева кокона солнечными лучами самки переворачивают его или перетаскивают с одного места на другое (Сычевская, 1935, 1945; Millot et Bourgin, 1942; Мариковский, 1956). Самки бродячих форм постоянно таскают кокон с собой, прикрепляя его к паутинным бородавкам (Lycosidae) или удерживая в когртках хелицер (Pisauridae). Так же поступают и некоторые виды пауков-тенетников, например Centromerus oviger Petr. и Lithyphantes oophorus Petr. (Petrunkevitch, 1926a).
Обычно "забота о потомстве" наблюдается только до вылупления молоди, но самки Lycosidae продолжают охранять молодых паучков и таскают их на брюшке. У Pardosa amentata С1. через 2-4 дня после отрождения молодые паучки покидают брюшко матери и расползаются (Vlijm et al., 1963). Забота о потомстве на этом заканчивается. У Coelotes terrestris Wid. молодь остается в гнезде матери 34 дня после выхода из кокона. В течение всего этого времени самка охраняет и кормит молодых паучков (Tretzel, 1961а, 1961b).
Коконы пауков обычно помещаются внутри гнезда, в котором также находится самка, охраняющая яйца. Кроме того, гнездо может служить убежищем для молодых паучков в первые дни их жизни. Гнезда пауков очень разнообразны. В одних случаях в качестве гнезда используется логовище, в котором паук живет постоянно. Так, нора пауков-норников (Lycosa, Alopecosa, многие Mygalomorphae) или логовище ловчей сети пауков-тенетников (некоторые Theridiidae, Araneidae и все Dysderidae) служат для самки гнездом во время откладки яиц и выхаживания молоди. В других случаях паук обычно не делает логовища, но в период размножения приготовляет специальное помещение для кокона, которое Вагнер (Wagner, 1894; Вагнер, 1910) называет "ложным гнездом". Гнезда этого типа характерны для многих Salticidae (рис. 107, 108). Самка, изготавливающая "ложное гнездо", как правило, не живет в нем.
Развитие. Яйца пауков относительно крупные, богатые желтком, покрытые двумя оболочками - желточной оболочкой и хорионом. Яйца оплодотворяются спермой, содержащейся в семеприемниках самки, во время яйцекладки. Самки пауков могут откладывать неоплодотворенные яйца, но лишь в редких случаях наблюдается развитие таких яиц (партеногенез). Факультативный партеногенез обнаружен у Filistata insidiatrix Forsk. (Damin, 1894), у Teutana triangulosa Walck. (Monterosso, 1944; Braun, 1956) и у Olios sericeus Kron. (Сычевская, 1962). Некоторые виды тропического семейства Ochyroceratidae, по-видимому, не имеют самцов и размножаются только нартеногенетически (Barros, 1964а, 1964b).
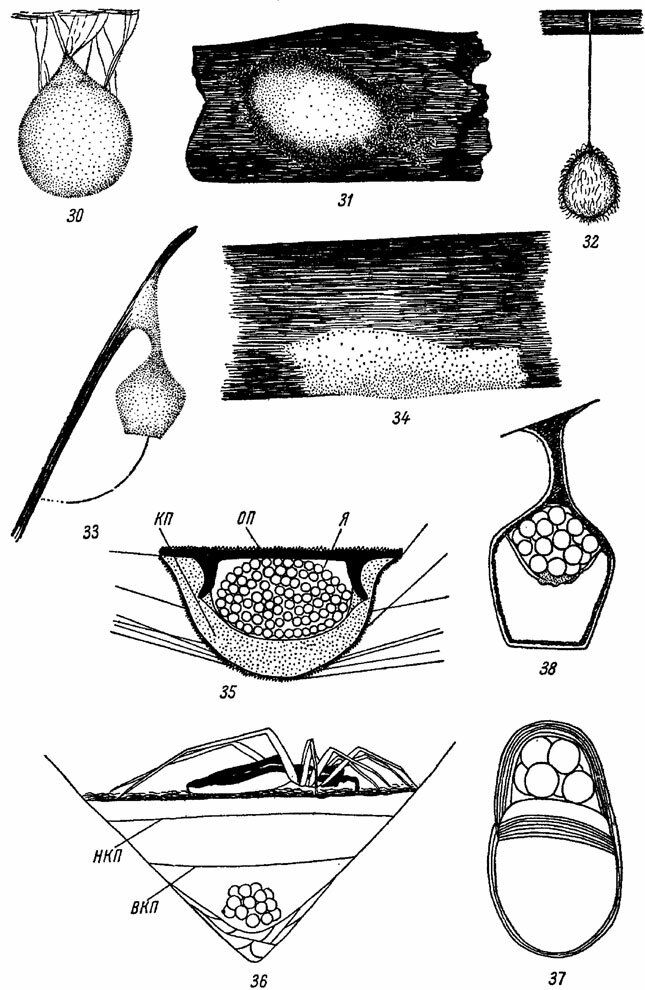
Рис. 30-38. Коконы пауков. По Мариковскому, Кастону, Хольму, Кроме и ориг. Рис. 30 Latrodectus tredecimguttatus Rossi (Tkeridiidae). Рис. 31. Agelena labyrinthica Cl. (Agelenidae). Рис. 32. Еrо furcata Vill. (Mimetidae). Рис. 33. Agroeca brunnea Black. (Clubionidae) Рис. 34. Tibellus oblongus Walck. (Thomisidae). Рис. 35. Araneus sp. (Агаneidae). Рис. 36. Philodromus dispar Walck. (Thomisidae). Рис. 37. Micaria pulicaria Sund. (Clubionidae) Рис. 38. Agroeca brunnea Black. BKП - внутренняя кроющая пластинка; КП - кроющая пластинка; НКП - наружная кроющая пластинка; ОП - основная пластинка; Я - яйца
Первая постэмбриональная стадия развития пауков резко отличается от всех остальных стадий и получает название пренимфы (Иванов, 1965). Пренимфа неспособна к движениям и не может питаться; тело ее докрыто эмбриональной кутикулой, не имеющей сочленовных мембран. Нимфы пауков довольно подвижны, но еще неспособны к самостоятельному питанию и к добыванию пищи. Нимфы во многом отличаются от взрослых пауков, так как у них недоразвиты глаза, ядовитые железы, паутинные органы, хелицеры и максиллы. Стадию пренимфы и нимфы пауки переживают в коконе. Пауки последующих преимагинальных стадий развития ведут самостоятельный образ жизни, активно питаются и отличаются от взрослых животных лишь недоразвитием половых органов. Имагинальный период развития - это период половозрелости. Обычно половозрелые пауки не линяют, но у Mygalomorphae линьки наблюдаются даже на имагинальной стадии развития (Buchli, 1960).
Линька пауков обычно протекает в две фазы. Вначале осуществляется отбрасывание щита головогруди и освобождение брюшка, и лишь затем происходит освобождение головогрудных придатков. Первая фаза линьки длится всего 30 минут, а вторая - несколько часов (Legendre, 1962). Общее число линек различно и зависит в первую очередь от размеров паука. Мелкие виды проделывают от 4 до 8 линек, а наиболее крупные - от 10 до 13. Самцы, как правило, имеют меньшее число преимагинальных возрастов и соответственно переживают меньше линек, чем самки.
Продолжительность развития пауков может колебаться от нескольких месяцев до нескольких лет. Особенно медленно развиваются крупные тропические виды пауков. Так, Aphonopelma hentzi требует для своего развития в зависимости от погодных условий от 8 до 13 лет. Взрослые самцы этого вида живут только 2-3 месяца, а взрослые самки - 10 лет и более (Baerg, 1963). Большинство видов европейской фауны успевает завершить весь цикл развития в течение летнего сезона. В лабораторных условиях при температуре 25° С днем и 15° С ночью развитие паука Zygiella x-notata Cl., продолжается около 3 месяцев (Le Guelte, 1963). Однако в природе этот вид развивается в течение 6 месяцев, и, следовательно, является моновольтинным видом, дающим лишь одну генерацию в году. По типу моновольтинных видов развиваются также пауки-крестовики Araneus folium Achrk., A. ocellatus Cl. (Бельтюкова, 1949), а также большинство других пауков умеренных широт. Поливольтинными видами, дающими 2-3 генерации в течение года, являются отдельные виды Linyphiidae и Micryphaniidae (Wiehle, 1956, 1960b; Heydemann, 1960), а также Araneus displicatus Hentz (Araneidae), Philodromus rufus Walck., Ph. aureolus Cl., (Thomisidae) и Paraphidippus marginatus Cby a. Bhp. из Salticidae (Dondale, 1961). Наконец, некоторые, главным образом южные по происхождению, виды пауков развиваются 2-3 года. Таковы Eresus niger Pet. (Norgaard, 1943a), Scytodes thoracica Latr. (Dabelow, 1958), Hyptiotes paradoxus C. L. Koch (Potzsch, 1963) и крупная раса тарантула Lycosa singoriensis Laxm. (Мариковский, 1956).
Зимой развитие пауков умеренных широт обычно приостанавливается, наступает диапауза. Однако пауки-синантропы развиваются круглый год (Бельтюкова, 1946), а отдельные виды очень мелких пауков сем. Micryphantidae спариваются и откладывают яйца исключительно зимой (Tretzel, 1954; Wiehle, 1960b; Broen u. Moritz, 1963). Субтропические и тропические пауки-норники рода Nemesia имеют летнюю диапаузу, которую они переживают в наглухо запечатанной норе (Buchli, 1960, 1961).
Зимуют пауки на самых различных стадиях развития. У большинства пауков-крестовиков рода Araneus зимуют яйца, а у Agelena labyrinthica Сl. и Zygiella x-notata Cl., - нимфы в коконе (Le Guelte, 1963; Potzsch, 1963). У Pardosa, Trochosa и других Lycosidae зимовка происходит в преимагинальной стадии и только у Alopecosa fabrilis Cl., из того же семейства - в стадии имаго (Wiebes, 1960). В тех случаях, когда развитие продолжается в течение 2 лет, зимуют различные преимагинальные стадии, как у Trochosa ruricola De Geer (Hackman, 1957, 1959), яйца и неполовозрелые паучки, как у Araneus diadematus Cl., (Wiehle, 1931a), или преимагинальные стадии и взрослые самки, как у Lycosa singoriensis Laxm. (Мариковский, 1956).
Экология (места обитания пауков). Места обитания пауков в природных условиях чрезвычайно разнообразны и включают в себя как пресные воды, так и самые различные наземные стации.
В пресных водах, преимущественно стоячих или полустоячих, обитает Argyroneta aquatica Cl., - единственный паук, который полностью приспособился к водному образу жизни. Этот паук дышит атмосферным кислородом, но постоянно живет под водой в специальном, наполненном воздухом паутинном колоколе, который прикрепляется к водным растениям. В таком колоколе осуществляется питание, спаривание и проходит все развитие паука (Сrоmе, 1951).
Многие Lycosidae и Pisauridae, обитающие по берегам рек, прудов и озер, прекрасно бегают по поверхностной пленке воды и даже могут входить под воду, свободно передвигаясь по дну. Среди подобных пресноводных пауков-амфибионтов наиболее известны Dolomedes fimbriatus Cl., Pardosa riparia С. L. Koch, Pirata hygrophilus Thor., P. piraticus Cl. Различные виды рода Arctosa также относятся к данной экологической группе и встречаются на влажном песке по берегам крупных стоячих водоемов или рек. Эти пауки, двигаясь по поверхности воды или по речному берегу, ориентируются по солнцу, определяя его положение по плоскости поляризации световых лучей (Papi, Serretti et Parrini, 1957; Tongiorgi, 1959, 1962; Papi et Tongiorgi, 1963).
Среди пауков также немало обитателей берегов океанов и морей. Часто такие пауки живут в зоне приливов и легко могут переносить продолжительное пребывание в морской воде без воздуха (Knulle, 1953; Wiehle, 1960а). В области литорали балтийского побережья ГДР аранеоценоз представлен 10-20 видами, причем доминирующим видом является Praestigia dujfeyi из сем. Micryphantidae (Heydemann, I960). Большинство пауков относится индифферентно к содержанию солей в почве, но пауки литорали, даже если они изредка и встречаются вдали от морских берегов, являются типичными галофилами и предпочитают наиболее засоленные участки биотопа (Hiebisch, 1962). Так, например, ареал паука-галофила Pardosa entzi Chyz. характеризуется чрезвычайной разорванностью и в настоящее время этот вид встречается по берегам морей и вблизи континентальных солоноватых водоемов (Tongiorgi, 1964). Он обнаружен, с одной стороны, на средиземноморском побережье Франции, Италии и Югославии, на о. Родос (Греция), на Южном берегу Крыма (СССР), а с другой стороны - на солончаках в Казахстане (СССР) (номинотипический подвид) и вблизи некоторых соленых озер Монголии (подвид estinensis).
Своеобразная фауна пауков складывается на болотах и заболоченных лугах, где преобладающее значение получают амфибионтные Lycosidae (Pardosa, Pirata и др.), а также пауки-тенетники сем. Tetragnathidae, являющиеся типичными гигрофилами. На влажных торфяных болотах доминируют различные Micryphantidae и Linyphiidae, многие из которых в своем распространении тесно связаны со сфагновыми мхами. Торфяные болота используются в качестве мест зимовки молодыми паучками семейств Thomisidae и Lycosidae (Kajak, 1959). По краям болот на листьях тростника (Phragmitis communis) в больших количествах встречается еще один паук-гигрофил clubiona phragmitis С. L. Koch.
Наземные пауки, составляющие подавляющее большинство отряда, в зависимости от способа ловли добычи, условий микроклимата и других факторов живут во многих разнообразных биотопах.
Пауки, обитающие в почве, сравнительно немногочисленны; одни из них заселяют различные углубления, полости и норы грызунов (Latrodeclus, Loxosceles, Palpimanus, Micaria, Sterrhochrotus), другие - строят самостоятельные норки (Lycosa, Eresus, Alopecosa, Atypus, Nemesia). Пауки - обитатели нор грызунов не являются специфическими нидиколами, а заползают в них в поисках пищи и убежища (Дубинин, 1946). Огромное число видов пауков встречается под камнями и на поверхности почвы. Наиболее характерны для этих биотопов Gnaphosidae (Zelotes, Gnaphosa и другие роды), Lycosidae (Pardosa, Alopecosa), Thomisidae (Oxyptila, Xysticus), Dysderidae (Dysdera), некоторые clubionidae и Agelenidae. На поверхности почвы часто встречаются также многие пауки-скакунчики (Salticidae), которые в период размножения забираются под камни, где их самки устраивают гнезда с коконами. В пустынях Средней Азии этих пауков часто можно видеть бегающими днем по раскаленной глинистой почве (Харитонов, 1948а).
Фауна лесной подстилки представлена многими видами из различных семейств. Обычно доминирующие виды лесной подстилки относятся к семействам Micryphantidae и Linyphiidae; так, в лесах окрестностей Осло (Норвегия) доминируют Cenlromerus arcanus Pick., Robertus scoiicus Jack., Asthenargus paganus Sim. (Palmgren, 1964). Однако зачастую здесь очень многочисленны также Gnaphosidae (Gnaphosa, Zelotes, Drassodes), мелкие Agelenidae (Cryphoeca, Hahnia), Lycosidae (Alopecosa, Pardosa, Trochosa) и некоторые Theridiidae. Общая численность пауков подстилки в среднем составляет от 50 до 150 экземпляров на 1 м2 (Schmidt, 1956), но сильно колеблется в зависимости от типа леса. Так, по данным В. Я. Шиперовича (1937, 1939), в Брянском лесном массиве на 1 м2 поверхности почвы приходится в сосново-дубовом лесу 31 паук, в сосняке-брусничнике - 70 пауков, а в сосновом лесу по болоту - 102 паука.
Травянистая растительность имеет свою обильную фауну пауков. Наиболее богато заселены пауками сложноцветные, зонтичные и лютиковые растения; сравнительно мало пауков на осоках и злаках. Только на цветках охотятся такие пауки, как Thomisus, Misumena, Synaema (Thomisidae). Менее прихотливы Xysficus, Tibellus из того же семейства, Oxyopidae и некоторые Salticidae, которые встречаются как на цветках, так и на листьях и стеблях растений. Виды родов clubiona и Chiracanthium (Clubionidae) очень часто живут на травянистых растениях, скрываясь в скрученных из листьев и опутаппых паутиной убежищах; во время размножения в подобных гнездах попадаются самки с коконами. Наконец, Dictynidae используют листья растений как площадку для постройки ловчих сетей. Иногда обнаруживается, что пауки отдают определенное предпочтение отдельным видам травянистых растений. Так, например, очень мелкий белый паучок Theridula ovsjannikovi Charit., распространенный в степях, живет только на Aster villosum и, может быть, также на Artemisia и Veronica (Харитонов, 1950).
Посевы сельскохозяйственных культур имеют сравнительно бедную аранеофауну, так как однообразная растительность, не образующая сомкнутого покрова, малоблагоприятна для целого ряда видов (Ажеганова, 1951). На посевах злаков доминируют различные Thomisidae и в первую очередь Xysticus ulmi Hahn; изредка здесь встречаются также Araneus adiantum Walck., Oxyopes lineatus Latr., O. heterophthalmus Latr. и Hypsosinga sanguinea C. L. Koch. В посевах люцерны наиболее обычны Раchygnatha degeeri Sund. в Средней Европе (Geiler, 1963), Oxyopes lineatus Latr. и Xysticus sp. в СССР на севере Узбекистана (Яхонтов, 1955).
Аранеофауна древесных пород и кустарников исключительно богата и разнообразна. В лиственных лесах преобладают Araneidae и Salticidae, в смешанных лесах заметно возрастает значение Theridiidae, хотя роль Araneidae еще довольно велика, а в хвойных лесах резко увеличивается численность Lycosidae, Linyphiidae, Thomisidae (Hansell, 1962). Надо отметить также, что видовой состав пауков изменяется не только при переходе от хвойных лесов к лиственным, но и в зависимости от типа леса (Харитонов, 1953). Это объясняется специфической приуроченностью отдельных видов пауков к определенным деревьям. Pachygnatha Clercki Sund., например, встречается почти исключительно на ольхе, a Zilla diodia Walck. - только на хвойных (Tretzel, 1954). Hyptiotes paradoxus С. L. Koch (Uloboridae) тоже поселяется исключительно на хвойных, явно предпочитая ели (Reukaf, 1931).
Ряд видов пауков являются специфическими обитателями стволов деревьев; одни из них встречаются на коре (Drapetisca socialis Sund., Philodromus emarginatus Schr., Salticus cingulatus Pan.), другие - только под корой (Coriarachne depressa С. L. Koch, Araneus umbraticus Cl.). Представители небольшого австралийского подсемейства крибеллятных пауков Matachiinae (Dictynidae) живут в стволах деревьев - в ходах насекомых, грызущих древесину (Marples, 1962а). Поздней осенью и зимой под отставшей корой деревьев скопляется большое количество различных пауков, находящих здесь убежище и часто устраивающих паутинные гнезда. Таковы некоторые виды рода clubiona, в частности С. corticalis Walck., С. pallidula Cl., и др.
Кроны деревьев имеют свою фауну пауков, которые в основном принадлежат к сем. Araneidae. Крупные пауки-крестовики рода Araneus раскидывают свои обширные сети между ветками деревьев; некоторые из них устраивают из листьев своеобразное убежище в стороне от ловчей сети (рис. 541). Более мелкие роды этого семейства (Cyclosa, Singa, Cercidia) часто тоже заселяют кроны. Кроме Araneidae для крон деревьев и для листвы кустарников очень характерно сем. clubionidae, представители которого обитают в свернутых и скрепленных паутиной листьях. Кроме того, на листве и ветках деревьев охотятся за насекомыми пауки-бокоходы (Philodromus, Xysticus) и пауки-скакунчики (Heliophanus, Dendryphantes).
Совершенно своеобразная группа пауков встречается в гнездах муравьев и термитов. К обитанию в гнездах муравьев приспособились представители совершенно различных семейств: Thyreostenius biouatus из Micryphantidae (Donisthorpe, 1927; Wiehle, 1960b), Myrmeques attarum Roew. из clubionidae (Wiehle, 1954), Bucranium sp. из Thomisidae (Bristowe, 1939-1941) и Zodarium vlasovi Sytsch. из Zodariidae (Власов и Сычевская, 1937). С гнездами термитов связаны почти исключительно представители сем. Oonopidae (Bristowe, 1939-1941; Benoit, 1964).
Многие пауки из различных семейств заселяют пещеры и гроты. В крымских пещерах, например, обнаружены Tegenaria taurica Charit. из Agelenidae, Meta bourneti Sim. и M. merianae Scop, из Araneidae, Leptyphantes knobarum Charit. из Linyphiidae, Pholcus phalangoides Fuess. из Pholcidae (Харитонов, 1947).
Почти все перечисленные виды пауков - это типичные ботриофилы, которые обитают исключительно в пещерах, и только P. phalangoides селится обычно в темных углах домов, в подвалах и погребах. Пещерный образ жизни ведет к исчезновению окраски покровов и недоразвитию глаз, что особенно хорошо заметно при сравнении открытоживущих и пещерных видов одного и того же рода. Так, темноокрашенные Porrhomma pygmaeum Blackw. (Linyphiidae) живут на почве и имеют нормально развитые, функционирующие глаза, а встречающиеся только в пещерах P. rosenhaueri L. Koch окрашены бледно и обладают сильно редуцированными глазами (Wiehle, 1956). Крайний случай редукции глаз наблюдается у Stalita taenaria Schiod., у которого атрофируются также зрительные нервы и оптические ганглии (Гааке, 1902).
Число известных видов пауков-синантропов достигает нескольких десятков. В домах и надворных постройках обитают некоторые представители сем. Theridiidae (Teutana castanea Cl., Т. triangulosa Walck.), все виды рода Pholcus и настоящие "домовые" пауки - Tegenaria domestica Cl., и T. ferruginea Panz. (Agelenidae). В южных районах СССР фауна синантропных видов пауков обогащается главным образом за счет сем. Pholcidae (Artema transcaspica Spass., Ceratopholcus maculipes Spass., Holocnemus pluchei Scop.). К ним присоединяются Uroctea nadiae Spass. (Urocteidae), Filistata crosbyi Spass. (Filistatidae) и Scytodes strandi Spass. (Sicariidae). Различные Oonopidae часто поселяются в домах и живут между страницами книг, где они охотятся за различными мелкими насекомыми (кожеедами, коллемболами и др.).
Питание. Все пауки - хищники, причем почти всегда они питаются только живой добычей. Пойманная добыча переваривается с помощью изливаемого на нее пищеварительного сока, обладающего сильным протеолитическим действием. Затем паук всасывает полупереваренную разжиженную пищевую массу. Состав добычи чрезвычайно разнообразен и включает в себя как мелких позвоночных, так и самых различных беспозвоночных животных.
Позвоночные животные, впрочем, очень редко служат добычей пауков, в особенности в пределах европейской части СССР. Амфибионтные пауки Dolomedes fimbriate Cl., D. plantarius Cl., иногда истребляют головастиков и мальков рыб (Харитонов, 1940; Schmidt, 1957), но более интересен в этом отношении Diaрontia oblongus C. L. Koch (Lycosidae) из Южной Америки, для которого мелкие рыбки служат, по-видимому, основной добычей (Berg, 1883). В литературе также отмечены факты нападения громадных тропических Mygalomorphae (сем. Aviculariidae) на амфибий, рептилий, птиц и даже на мышевидных млекопитающих (Millot, 1943). Веллард (Vellard, 1936), например, в своей монографии приводит великолепную фотографию бразильского паука, атакующего небольшую змею, а П. И. Мариковский (1956) описывает случай использования в пищу тарантулом (Lycosa singoriensis Laxm.) молодой ящерицы.
Подавляющее большинство пауков питается беспозвоночными животными, главным образом насекомыми. Набор видов насекомых, которые используются в пищу теми или другими пауками, зависит в первую очередь от способа добычи пищи (стр. 25), Так, среди жертв пауков-кругопрядов (сем. Araneidae), которые ловят насекомых с помощью крупных колесовидных сетей, 25.7% составляют Diptera, 18.4% - Trichoptera, 11.2% - Neuroptera, 10.5% - Hymenoptera и 9.9% - Lepidoptera (Turnbull, 1956). По данным П. Воронцовского (1931), Argiope lobata Pall, из этой же группы пауков является активным истребителем саранчовых. В питании других пауков-тенетников основное значение также имеют Diptera и Hymenoptera и в меньшей степени - некоторые Hemiptera и Homoptera. Несколько иной состав добычи у паука Agelena naevia Bosc. из Северной Америки, который подобно нашему A. labyrinthica Cl., строит воронковидную сеть в низкой траве, непосредственно над поверхностью почвы. По наблюдениям Бильзинга (Bilsing, 1920), в пище этого паука 55% составляют Orthoptera, 12% - Hymenoptera (муравьи), 9% - Auchenorhyncha и только 8% - Diptera. Мелкие пауки лесной подстилки и верхнего слоя почвы питаются в основном ногохвостками (Collembola); особенно часто представители этого отряда используются в пищу молодыми паучками сем. Micryphantidae (Bristowe, 1939-1941; Wiehle, 1960b). Отряд. Coleoptera составляет обычную добычу пауков-норников - тарантула Lycosa singoriensis Laxm. (Мариковский, 1956) и Eresus niger Pet. (Norgaard, 1943a). В основном на жуков охотится также паук Coelotes terrestris Wid. (Agelenidae), изготавливающий трубчатую сеть (Tretzel, 1961а). Обычно пауки нападают только на взрослых насекомых и значительно реже питаются личинками. Добыча Linyphia triangularis Cl., например, на 90% состоит из взрослых насекомых и лишь на 10% - из личинок (Turn-bull, 1960). Гусеницы особенно часто отвергаются пауками и лишь в исключительных случаях принимаются ими. Многие пауки также отказываются поедать жуков-нарывников, "божьих коровок", клопов (за исключением Miridae), жалоносных перепончатокрылых (за исключением муравьев), пилильщиков и целый ряд других насекомых (Bilsing, 1920; Bristowe, 1939-1941; Мариковский, 1956; Тыщенко, 1961).
Нередко пауки нападают кроме насекомых и на других наземных или водных беспозвоночных. Основную добычу паука-серебрянки (Argyroneta aquatica Cl.) составляют водяные ослики (Asellus aquaticus), но иногда он уничтожает и мелких моллюсков-(Poujade, 1888; Кудринская, 1955). Наземные ракообразные (класс Isopoda) используются в пищу представителями рода Latrodectus (L. mactans Fabr. - Exline a. Hatch, 1934; L. pallidus pavlovskyi Charit. - Уточкин, 1956) и средиземноморским пауком Uroctea durandi Walck. (Crome, 1957), Пауки могут питаться также многоножками и различными паукообразными: сенокосцами и клещами (Bristowe, 1939-1941), фалангами и скорпионами (Мариковский, 1956) или другими пауками (Bilsing, 1920; Bristowe, 1939-1941; Tretzel, 1961а, и др.). Очень распространен среди пауков каннибализм - поедание особей своего вида.
Специализация на строго определенной пище (монофагия) наблюдается крайне редко и свойственна паукам, питающимся муравьями или другими пауками. Так, Dipoena tristis Hahn и Theridium riparium Black, из Theridiidae нападают только на муравьев и никогда не трогают других насекомых (Wiehle, 1937; Vite, 1953; Kirchner, 1964). Виды рода Zodarium также являются типичными мирмекофагами (Simon, 1914; Власов и Сычевская, 1937; Wiehle, 1954). Пауки двух неродственных семейств (Mimetidae и Archaeidae) целиком специализировались на питании другими пауками и всегда отказываются от всякой иной добычи (Wiehle, 1953; Legendre, 1961; Czajka, 1963). Олигофагия представляет довольно обычное явление у пауков; примером олигофагии может служить уже отмеченное выше питание пауков-норников, которые предпочитают охотиться на различных жуков, но при случае не отказываются и от другой добычи. Наконец, очень многие пауки нападают практически на любое животное, которое могут осилить. В качестве иллюстрации можно привести состав пищи Uroctea durandi Walck. (Grome, 1957). Этот паук одинаково охотно схватывал и высасывал личинок и жуков мучного хрущака (Tenebrio), небольших жужелиц (Pterostichus, Harpalus), жуков-дровосеков (Leptura, Acanthocinus), мух (Musca, Fahnia, Calliphora), прямокрылых (Stenobothrus, Tettigonia), чешуйниц (Lepisma), мокриц (Oniscus, Роrcellio) и молодых многоножек (Schizophyllum).
Все пауки охотно пьют воду. При наличии воды они в состоянии голодать очень долго, обычно в течение нескольких месяцев. Так, например, установлено, что самки Steatoda bipunctata L. могут 1.5 года прожить без пищи (Comstock, 1912).
Способы добычи пищи. Анцестральные пауки, по-видимому, были бродячими охотниками и пользовались паутиной только для изготовления коконов (Kaston, 1964). Среди современных пауков наиболее близки к этим предполагаемым анцестральным формам многие представители семейств Lycosidae и Pisauridae, которые в быстром беге настигают свою добычу. Pardosa amentata С1., например, ведет дневную бродячую жизнь, не строит постоянного убежища и активно преследует любую добычу, которая попадает в его поле зрения (Dahl, 1908; Vlijm, Kessler et al., 1963).
Однако уже среди Lycosidae многие виды переходят к оседлой жизни и начинают строить логовища в виде земляных норок. Довольно глубокие вертикальные норы изготовляют тарантулы Lycosa singoriensis Laxm., L. narbonensis Walck, и другие виды рода Lycosa (Вагнер, 1886а; Каландадзе и Мхеидзе, 1955; Мариковский, 1956; Crome, 1956; Kuenzler, 1958). В целях защиты от врагов тарантул иногда снабжает входное отверстие норки паутинной закраиной или небольшим колпачком; L. marikovskii окружает устье норки высокой трубочкой из частиц почвы, скрепленных паутиной, a Alopecosa opifex Wag. даже делает норки с захлопывающейся крышечкой-дверцей (Иванов, 1965). Во всех этих случаях пауки уже не бродят в поисках добычи, а поджидают ее, укрываясь в норе.
Подобный способ охоты за добычей с использованием норы-убежища широко распространен и среди других семейств пауков. Весьма совершенные земляные норки, часто снабженные шарнирными крышечками, строят Ctenizidae, Migidae, Barychelidae и Eresidae. У Atypus паутинная выстилка подземной норки продолжается на поверхности земли в виде замкнутой цилиндрической трубки, внутри которой сидит паук, поджидающий добычу (Simon, 1874; Ehlers, 1937; Спасский и Миненкова, 1940; Wiehle, 1953; Mikulska, 1954; Gray, 1956; Blazejewski, 1958; Hallander, 1958; Ressl, 1960; Broen u. Moritz, 1964). Когда по этой надземной трубке пробегает какое-нибудь насекомое, паук захватывает его прямо сквозь паутинную ткань. Поврежденная при этом стенка трубки вскоре восстанавливается.
Различными способами подстерегания добычи на цветках, на листьях, на коре деревьев или на скалах пользуются пауки из семейств Thomisidae, Sparassidae и clubionidae. Эти пауки нередко обладают превосходной покровительственной окраской, облегчающей охоту за различными насекомыми (см. стр. 6), Мадагаскарские пауки сем. Archaeidae питаются исключительно другими пауками и подкарауливают проходящую мимо жертву, сидя в засаде (Legendre, 1961). С другой стороны, европейский Еrо furcata Vil. (Mimetidae) охотится за самками паука-кругопряда Meta segmentata Cl., имитируя поведение самца этого вида (Czajka, 1963). Наконец, тропические виды рода Phrynarachne из Thomisidae замечательно точно имитируют помет птиц на листьях и даже издают запах птичьих экскрементов (Jacobson, 1921). Этот запах привлекает некоторых мелких мух, которые и составляют добычу паука.
Salticidae. и Oxyopidae медленно подкрадываются к добыче и завладевают ею одним прыжком. Обладая хорошим зрением и подвижными трубкообразными передними медиальными глазами, пауки сем. Salticidae могут замечать насекомое издалека и точно оценивать расстояние до него (Dzimirski, 1959). Так, например, Evarcha замечает движущуюся муху уже на расстоянии 20-25 см (Homann, 1928). Когда муха приближается на расстояние в 4 см, паук подкрадывается к ней и, наконец, на расстоянии в 1.5 см стремительно прыгает на добычу.
Некоторые пауки не строят специальной ловчей сети, но тем не менее при ловле добычи пользуются паутинными нитями. Средиземноморский паук Uroctea durandi Walck. ловит добычу возле своего логовища, забрасывая жертву длинными и широкими лентами из клейкой паутины (Crome, 1957). Некоторые тропические пауки охотятся с помощью одиночной клейкой нити, которую они набрасывают наподобие аркана на приближающееся насекомое и затем подтягивают к себе приклеившуюся добычу (Gertsch, 1947, 1955; Kaston, 1965). Такой своеобразный способ ловли добычи был описан для Mastophora из Америки (Hutchinson, 1903), для Dicrostichus из Австралии (Longman, 1922) и для Cladomelea из Южной Африки (Akerman, 1923). В последнем случае паук удерживает "аркан" III и IV парами ног и быстро раскручивает его над собой в горизонтальной плоскости. У Mastophora и Dicrostichus "аркан" удерживается передними ногами и не раскручивается.
Оригинальный способ добычи пищи используют пауки рода Scytodes из сем. Sicariidae (Monterosso, 1927; Dabelow, 1958; McAlister, 1961), Пауки этого рода обладают очень крупными ядовитыми железами, которые в своей задней части преобразованы в паутинные железы, выделяющие клейкий секрет (Millot, 1929, 1949). Заметив добычу на расстоянии 1-2 см, Scytodes опрыскивает ее выделениями этих желез. Жидкий декрет быстро затвердевает в виде зигзагообразных паутинных нитей, прочно приклеивающих жертву к субстрату (рис. 99).
Простейшую паутинную нить, связанную с норкой, которая служит убежищем паука, строят Liphistiomorphae, некоторые Mygalomorphae, Eresidae, Filistatidae и Amaurobiidae из Cribellatae, а также Dysderidae из Ecribellatae. У Filistata и Атаиrоbins (рис. 39) сеть состоит из паутинных нитей, которые радиально расходятся от входа норки (Comstock, 1912; Norgaard, 1951; Wiehle, 1953; Cloudsley-Thompson, 1955). Одни из этих нитей поддерживают край паутинной выстилки убежища, другие играют роль сигнальных нитей, предупреждающих паука о приближении добычи, или даже ловчих крибеллятных нитей, снабженных клейким веществом. Пауки при ловле добычи располагаются у самого входа в норку; когда добыча коснется сигнальных нитей, связанных с убежищем, они быстро выбегают наружу, хватают ее и снова скрываются в норке.
Даже у пауков перечисленных выше семейств очень часто земляная норка заменяется паутинной трубкой. Однакд высшую ступень развития сетей этого типа составляют постройки Agelenidae, где устье трубки расширяется в виде воронки, связанной с обширной ловчей сетью типа тента или полотнища (рис. 40). Такую воронко-видную ловчую сеть строит, например, наш обычный паук Agelena labyrinthica Cl., (Спасский, 1920; Schlott, 1931; Wiehle, 1949). Паук сидит у входа в трубку-убежище и подстерегает насекомых, заползающих на ловчую сеть. У другого представителя этого же семейства - Coelotes terrestris Wid. изготавливается U-образная паутинная трубка, к обоим концам которой примыкает ловчая сеть (Tretzel, 1961а). В этом случае паук также сторожит добычу у входа в паутинную трубку.
Крибеллятные пауки сем. Dictynidae изготовляют ловчую сеть, тесно связанную с убежищем из паутины. При этом Ergatis walckenaeri Roew. располагает клейкие ловчие нити на поверхности древесного листа, a Dictyna arundinacea L. протягивает их из убежища, расположенного в развилке двух сухих веточек, к соседним ветвям дерева (Wiehle, 1949, 1953). Dictyna civica Luc. делает плоские ловчие сети и поселяется, часто в огромных количествах, на стенах домов (Billaudelle, 1957).
Сравнительно простую беспорядочную сеть строят некоторые Theridiidae. Однако уже у Teutana, Steatoda, Lithyphantes и у многих Theridium существует определенный порядок в расположении отдельных элементов ловчей сети (рис. 41, 42). У этих пауков сеть имеет горизонтальный полог из редкого сплетения простых паутинных нитей и вертикальные клейкие нити, образующие ловушку для муравьев и других ползающих насекомых (Nielsen, 1928-1932; Wiehle, 1931b, 1937, 1949; Holm, 1939; Norgaard, 1948, 1956). У Theridium riparium Blackw. (рис. 42) сеть строится непосредственно над землей (не выше 11 см) и снабжается убежищем в виде колпачка, обклеенного снаружи кусочками почвы (Norgaard, 1956; Freisling, 1961). Сети пауков рода Latrodectus (рис. 354) также состоят из логовища и собственно ловчей сети, которая имеет горизонтальную платформу и тонкие вертикальные клейкие нити (Мариковский, 1956; Уточкин, 1956; Szlep, 1965).
Мелкие пауки сем. Theridiidae иногда поселяются в сетях крупных крестовиков и питаются остатками жертв хозяина. Широко распространенный по всему земному шару паук Theridium tepidariorum С. L. Koch обычно устраивает свою сеть на периферии паутины Cyrtophora citricola For. и может пользоваться добычей, попавшей в сеть хозяина (Kullmann, 1960а, 1960b). Виды рода Argyrodes - облигатные комменсалы, которые живут в сетях Cyrtophora или других крупных пауков, питаясь на пойманной ими жертве и оставаясь незамеченными хозяином благодаря своим малым размерам (Exline a. Levi, 1962; Tikader, 1963). Истинными комменсалами являются также все известные виды рода Conopistha, которые обитают в сетях различных Araneidae, Linyphiidae, Agelenidae, Pholcidae и Theridiidae (Exline, 1945; Thomas, 1953; Lamore, 1957; Kullmann, 1959; Kaston, 1965).
Крышеобразные или куполообразные ловчие сети делают представители семейств Linyphiidae, Pholcidae и Hypochilidae. У Linyphia triangularis Cl., и у других Linyphiidae сеть имеет плотный горизонтальный тент и отходящие от него вверх и вниз поддерживающие нити (Nielsen, 1928-1932; Norgaard, 1943b; Wiehle, 1949, 1956; Turn-bull, 1960). Насекомые, пролетающие над такой сетью, ударяются о верхние поддерживающие нити и падают на горизонтальный тент. Паук во время охоты сидит с нижней стороны тента и хватает падающую добычу, прорывая хелицерами плотную паутинную ткань его. У Lepthyphantes obscurus Blackw. в отличие от других видов семейства паук, поджидающий добычу, сидит головой вниз на стволе дерева, а тент сети располагается не горизонтально, а вертикально (Kullmann, 1961b). Drapetisca socialis Sund., который также охотится на стволах деревьев, строит сильно упрощенную неправильную сеть, предназначенную лишь для сигнализации о добыче (Kullmann, 1961а). Многие Micryphantidae, морфологически очень близкие к Linyphiidae. также имеют вторично упрощенные крышеобразные ловчие сети (Wiehle, 1960b).
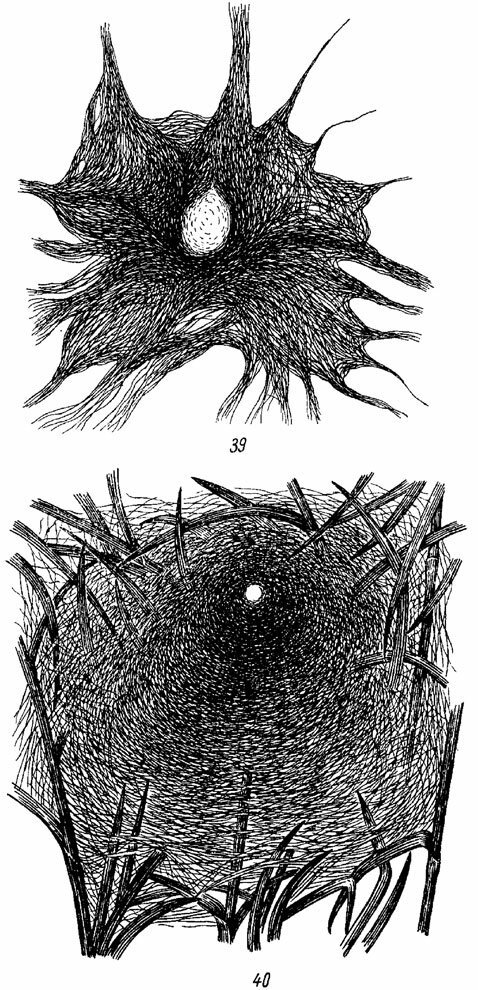
Рис. 39, 40. Ловчие сети пауков. Ориг. Рис. 39. Amaurobius sp. (Amaurobiidae). Рис. 40. Agelena labyrhinthica (Agelenidae)
Наиболее совершенными сетями пауков бесспорно будут колесовидные сети, в которых достигается максимальная эффективная поверхность ловчей зоны при минимальных затратах паутины. Такие сети обнаруживаются только в четырех семействах пауков - Uloboridae, Araneidae, Tetragnathidae и Tkeridiosomatidae.
Сеть Araneus diadematus Cl., (рис. 537) состоит из наружной рамы, к которой крепятся радиусы, отходящие от центральной сеточки (Berland, 1932; Peters, 1933а; Gerhardt u. Kaestner, 1937-1938; Иванов, 1946, 1965; Wiehle, 1949; Харитонов, 1953; Breed et al, 1964; Grasshoff, 1964; Reed et al., 1965; Witt, 1965, и др.). Ловчая зона такой сети образуется из многочисленных спиральных оборотов единственной клейкой нити; эта зона отделяется от центральной сеточки свободной зоной. У других видов рода Araneus (например, у A. marmoreus Gl. - рис. 541) сеть имеет аналогичное строение, но иногда снабжается убежищем, вынесенным на периферию или за пределы сети и соединенным с нею сигнальной нитью (Wiehle, 1928, 1931 а). У Zygiella (рис. 539) сигнальная нить проходит в пустом секторе, лишенном клейких ловчих нитей (Wiehle, 1931а, 1949; Braun, 1955; Le Guelte, 1965; Witt, 1965). Сети некоторых Araneidae снабжены так называемыми стабилиментами (рис. 538), которые представляют собой широкие прямые или зигзагообразные иногда пушистые ленты из густой белой паутины (McCook, 1889-1892; Tilquin, 1942; Millot, 1949; Zollinger, 1955; Tikader. 1961). Стабилимент, по-видимому, служит для маскировки паука и встречается у тех видов, которые всю жизнь проводят в центре сети.
Очень тонкая и практически невидимая ловчая сеть пауков-кругопрядов представляет собой прекрасную ловушку для летающих насекомых. Клейкие нити ловчей зоны прочно удерживают крылья попавших в сеть двукрылых, перепончатокрылых, саранчовых и стрекоз; однако колесовидная сеть менее пригодна для ловли бабочек и ручейников, крылья которых покрыты волосками или чешуйками (Eisner et al., 1964). Радиальная конструкция ловчей сети создает возможность распространения всех возникающих в ней вибраций по направлению к центральной сеточке, где обычно сидит паук во время охоты. Определение местонахождения добычи в пределах сети производится пауком на основании этих вибраций (Peters, 1931, 1933b; Liesenfeld, 1956, 1961; Szlep, 1964). Однако в начальной фазе захвата добычи у Araneus diadematus имеет значение не только вибрационное, но и оптическое раздражение (Giulio, 1962). Направление движения паука, по-видимому, определяется совместным действием обоих раздражителей.
Ловчая сеть пауков-кругопрядов обычно имеет строго постоянную структуру: постоянное количество радиусов и витков, спиральной клейкой нити, постоянное расстояние между двумя соседними витками и т. д. Так, в сети взрослой самки Araneus diadematus имеется 39 радиусов, 35 витков спирали и 1225 точек прикрепления радиусов к спиралям (Witt, 1965). Сети неполовозрелых форм этого же паука сохраняют типичный для вида общий план строения, но имеют увеличенное число радиусов и витков спирали (Witt, Baum, 1960). Типичную круговую паутину молодые паучки начинают плести уже через 2 недели после выхода из яиц, даже если они никогда не видели ловчей сети и старых пауков (Witt, 1963). Очевидно, определенный порядок в расположении и числе основных элементов ловчей сети представляет собой результат запрограммированного наследственностью автоматического поведения паука при постройке сети.
При постройке сети паук-крестовик Araneus diadematus первоначально изготовляет трех- или четырехугольную раму и затем протягивает радиусы из центра наружу (Berland, 1932; Peters, 1953; Иванов, 1965). Каждый раз возвращаясь в центр, паук заплетает центральную сеточку и расположенную вокруг нее прикрепительную зону. Спиральная нить изготавливается в два приема: первоначально паук протягивает вспомогательную временную спираль из нескольких оборотов сухой паутины, а затем, двигаясь от периферии к центру по этой временной спирали, животное проводит дефинитивную частую спираль из клейких ловчих нитей. Временная спираль при этом постепенно уничтожается. Через каждые 1-2 дня пауки разрушают паутину и ткут новую (Breed et al., 1964).
Строго постоянные соотношения между размерами отдельных частей сети у разных видов пауков-кругопрядов можно было бы объяснить либо постоянным соотношением длины ног паука, либо регулирующим влиянием центральной нервной системы. Первое из этих объяснений, по-видимому, неудовлетворительно, так как, например, у Zygiella x-notata Cl., передние ноги по отношению к другим ногам становятся длиннее с каждой последующей линькой, а соотношения частей ловчей сети остаются одинаковыми у пауков всех возрастов (Peters, 1954; Le Guelte, 1965). Второе объяснение подтверждается результатами опытов с введением паукам наркотиков или фармакологических агентов, действующих на нервную систему; в этих опытах наблюдалось сильное изменение структуры ловчей сети после инъекции препарата (Peters u. Witt, 1948; Peters, 1954; Witt, 1954, 1956; Christiansen et al, 1962: Witt a. Reed, 1965).
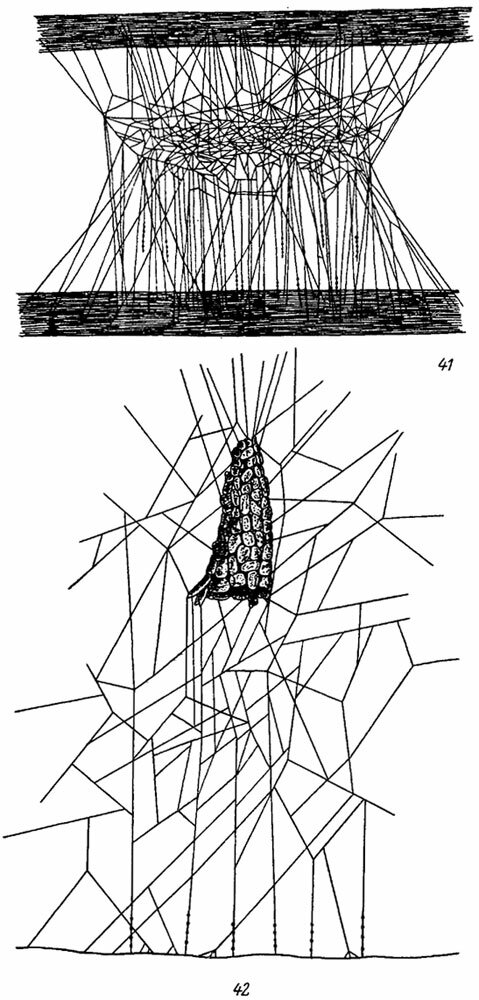
Рис. 41, 42. Ловчие сети пауков сем. Theridiidae. По Виде. Рис. 41. Steatoda sp. Рис. 42. Theridium riparium Black
Ловчие сети Tetragnathidae имеют очень много общего с уже рассмотренными сетями Araneidae, но характеризуются отсутствием центральной сеточки. Впрочем, подобный "открытый центр" обнаруживается и у некоторых Araneidae (например, у Meia). Сеть Eucta kaestneri Сr. из Тetragnathidae имеет сравнительно небольшое количество (10-15) радиусов и только 8-10 витков ловчей спирали (Crome, 1954). Сеть располагается на высоте 70 см над землей, а основную добычу паука составляют Nematocera, главным образом Tendepedidae и Culicidae. Подобные ловчие сети строят и различные виды рода Tetragnatha.
Типичную круговую сеть изготовляет Theridiosoma gemmosum L. Koch (Theridiosomatidae, рис. 44). Длинная сигнальная нить тянется от середины сети и прикрепляется к ближайшему растению. Паук, располагаясь возле центра сети, так сильнонатягивает сигнальную нить, что вся сеть приобретает форму конусовидного мешка. (McCook, 1889-1892; Wiehle, 1929, 1931а). Как только насекомое коснется ловчей сети, паук отпускает сигнальную нить и жертва запутывается в клейких ловчих нитях.
Колесовидная ловчая сеть крибеллятных пауков из сем. Uloboridae, по-видимому, возникла независимо от похожих на нее сетей Araneidae (Szlep, 1961; Kaston, 1964). У Uloborus сеть имеет в основном такую же форму, как у Araneus, но располагается горизонтально и часто снабжается стабилиментом (Marples, 1962а). У Hyptiotes paradoxus С. L. Koch (рис. 43) паук делает только один сектор типичной круговой сети с длинной сигнальной нитью, которая удерживается пауком (Reukaf, 1931; Wiehle, 1931b). Когда насекомое пролетает через такую сеть, паук слегка отпускает сигнальную нить и захлестывает его клейкими нитями из "крибеллятной" паутины.
Представители тропического семейства крибеллятных пауков Dinopidae ловят добычу с помощью сильно видоизмененной круговой сети. Наблюдения над африканским Menneus camelus Рос. (Akerman, 1926), австралийским Dinopis subrufa L. Koch (Roberts, 1954a, 1955) и американским D. spinosa Marx (Theuer, 1954) показали, что все пауки этого семейства делают небольшую ловчую сеть в виде треугольной или четырехугольной рамочки с натянутыми на нее частыми и параллельными клейкими нитями. Готовая сеть во время охоты удерживается передними ногами паука. Когда приближается насекомое, паук, раздвигая передние ноги, растягивает эластичную сеточку и опутывает ею добычу.
Среди Uloboridae сеть Hyptiotes представляет собой первый, начальный этап редукции полной круговой сети, достигающей своего крайнего выражения у Miagrammopes из Южной Африки (Akerman, 1932; Wiehle, 1954; Marples, 1962а, 1962b). Этот паук изготовляет только одну горизонтальную нить длиной от 0.9 до 3.6 м; средняя часть этой нити покрывается клейкой крибеллятной паутиной. Прикрепляя один конец подобной линейной "ловчей сети" к ветке дерева, паук поддерживает ее в растянутом состоянии. Как только насекомое задевает нить, паук мгновенно ослабляет ее, запутывая добычу.
Вторичное упрощение сетей до одной или нескольких нитей наблюдается и в некоторых семействах некрибеллятных пауков. У европейских видов Episinus из Theridiidae в ловчей сети сохраняются всего 2 клейких вертикальных нити, которые посередине соединяются поперечной сухой паутинкой, образуя Н-образную конструкцию (Holm, 1939). Паук сидит на поперечной перекладине сети, а свои передние ноги располагает вдоль одной из вертикальных нитей, У обитающего в Новой Зеландии паука Elesanis из того же семейства описана линейная сеть из одиночной нити длиной около 10 см; положение паука при охоте и способы ловли добычи здесь такие же, как у Miagrammopes (Marples, 1955), Наконец, в отдельных случаях (например, у Pachygnatha из Тetragnathidae) способность к постройке ловчих сетей у взрослых пауков полностью утрачивается и сохраняется только в молодых предымагинальных возрастах (Balogh, 1934).
Враги. Среди позвоночных животных наиболее существенные враги пауков - это насекомоядные птицы, которые особенно часто используют их для выкармливания птенцов. Дрофа (Otis torda) поедает каракуртов в таких количествах, что желудок этих птиц бывает набит ими (Констансов, 1907). По наблюдениям М. Н. Ионова (1948), боялычная соня (Selevenia betpakdalensis) питается различными паукообразными и в неволе охотно поедает тарантулов. Отмечается также истребление пауков мелкими млекопитающими, ящерицами, змеями и амфибиями.
Пауки иногда входят в состав добычи некоторых хищных насекомых - складчатокрылых ос (Vespidae), мух-ктырей (Asilidae), богомолов (Mantidae). Жуки-бронзовки Pseudospilophorus plagosus поедают пауков Stegodyphus mimosarum Pav, которые искусственно разводятся в Южной Африке для борьбы с комнатной мухой. Медведка (Gryllotalpa unispina) истребляет тарантулов ранней весной, когда пауки слабо подвижны, вялы и почти неядовиты; однако летом она сама становится добычей тарантула (Мариковский, 1956).
Многие осы-парализаторы из семейств Pompilidae и Sphecidae охотятся за пауками и обездвиживают свою жертву уколом жала в нервные ганглии. Такие парализованные, неподвижные пауки еще долго остаются живыми и используются осами для выкармливания личинок. Для каждой личинки обычно заготавливается только одна жертва, размеры которой превосходят размеры осы. Охотничьи приемы ос-парализаторов различны в соответствии с различной экологией и поведением их жертв. Так, Episyron tripunctatus из Pompilidae охотится на крестовика Araneus sp., легко передвигаясь по клейким нитям круговой ловчей сети (Berland, 1958). Почуяв приближение врага, паук бросается вниз, повисая на паутинной нити, по которой его и настигает Оса. Виды рода Pepsis, которые охотятся за крупными тропическими пауками Aphonopelma, зачастую вообще не подходят вплотную к пауку, а парализуют его на некотором расстоянии, изогнув и вытянув брюшко возможно дальше вперед (Petrunkevitch, 1926b; Williams, 1956; Cazier a. Mortenson, 1964). Особенно интересны повадки тех ос-помпилов, которые парализуют пауков сем. Ctenizidae, обитающих в глубоких земляных норках с шарнирной крышечкой. Эти осы легко открывают крышечку норы и смело бросаются в логовище паука (Millot, 1949).
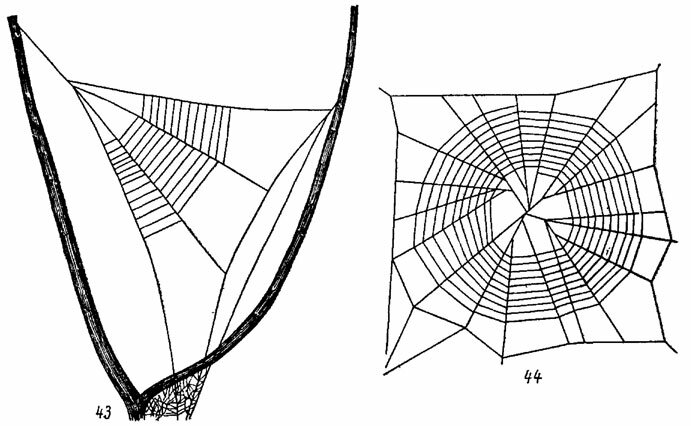
Рис. 43, 44. Ловчие сети пауков. По Виле и ориг. Рис. 43. Hyptiotes paradoxus С. L. Koch (Uloboridae). Рис. 44. Theridiosoma gemmosum L. Koch (Theridiosomatidae)
Сложное инстинктивное поведение парализующей осы во время охоты обусловливает строгую специализацию этих насекомых в отношении их добычи. Поэтому многие осы-помпилы нападают только на определенный вид или род пауков. Установлено, например, что Anoplius fulgidus в США нападает лишь на амфибиотического паука Pirata sedentaria (Wasbauer, 1955), a A. imbellus, которая встречается в той же местности, парализует только паука Pardosa, ramulosa (Wasbauer, 1957), В Финляндии добычу помнила Deuteragenia intermedia составляют исключительно пауки сем. Thomisidae, главным образом Xysticus audax Schr. (Valkeila, 1955), а в СССР в Средней Азии Pseudagenia rufiventris нападает на Olios sericeus Cron. и лишь в редких случаях - на Oxyopes lineatus Latr. (Сычевская, 1962).
Весьма многочисленны паразиты пауков. Взрослые пауки и нимфы иногда бывают заражены нематодами Mermis и Gordius или эндопаразитическими личинками мух из сем. Cyrtidae (Millot, 1949). Однако гораздо чаще паразитические насекомые, в первую очередь различные наездники (Ichneumonidae, Chaleididae), мухи (Tachinidae, Chloropidae) и сетчатокрылые (Mantispidae), развиваются за счет яиц пауков. Наездник Tromatobia ovivora, например, откладывает яйца в коконы пауков-крестовиков Araneus quadratus Cl., и A. diadematus Cl., (Crome, 1959). В один кокон паразит откладывает 4-8 яиц, но развивается только одна личинка, которая уничтожает до 75% яиц паука, затем окукливается и вылетает на следующий год. По данным Е. А. Волянской (1960), этот же наездник, а также Tromatobia concors заражают коконы каракурта. П. И. Мариковским (1956) приводится еще три вида наездников, паразитирующих на яйцах каракурта. Наиболее опасным врагом паука автор считает наездника Gelis marikovskii, который отличается быстрым темпом развития и может заражать также коконы Lycosa singoriensis Laxm., Agelena labyrinthica Cl., и Lithyphantes paykullianusWalck. (Мариковский, 1956). Коконы американского вида Latrodectus mactans Fabv., близкого к нашему каракурту, заражаются не только наездниками, но и мухами Pseudogaurax signatus из сем. Chloropidae (Jenks, 1938),
клетка для хомяка купить выгодно тут
|
ПОИСК:
|

При использовании материалов сайта активная ссылка обязательна:
http://paukoobraznye.ru/ 'Паукообразные: клещи, пауки, скорпионы'